在功能上,真核生物基因组分为转录活跃的常染色质和转录沉默的异染色质。在拟南芥中, 常染色质位于含有大多数转录活性基因的染色体臂上,其表达取决于组织/细胞类型、发育阶段和环境条件。而异染色质缺乏蛋白质编码基因,通常与端粒、着丝粒、转座子、沉默的 rDNA和散布在常染色臂上的小的异染色质岛相关。多种分子途径的相互作用可以调节异染色质状态,包括DNA甲基化、组蛋白修饰和RNA沉默。在拟南芥中,CHG和CHH甲基化主要与异染色质相关,并在功能上与H3K9位点的甲基化相关。另一个关键的抑制性组蛋白标记是H3k27甲基化(H3K27me),它独立于DNA甲基化途径。多梳抑制复合物1(PRC1)是一类重要的表观遗传调控因子, 它具有H3K27me3-reader和组蛋白H2A E3泛素连接酶的活性,并通过组蛋白H2A赖氨酸119单泛素化(H2AK119ub1)介导基因的抑制。在招募PRC1到染色质上出现了几个因素,如最近出现的同源异型蛋白转录因子NDX(NODULIN HOMEOBOX),它是迄今为止唯一与PRC1相互作用的转录因子。
近日,匈牙利的科学家Lóránt Székvölgyi在国际著名杂志Nature Communications在线发表了“NODULIN HOMEOBOX is required for heterochromatin homeostasis in Arabidopsis”的研究性论文。他们使用细胞学、分子生物学和高通量测序的方法描述了NDX基因在基于染色质的基因调控过程中的全局作用。
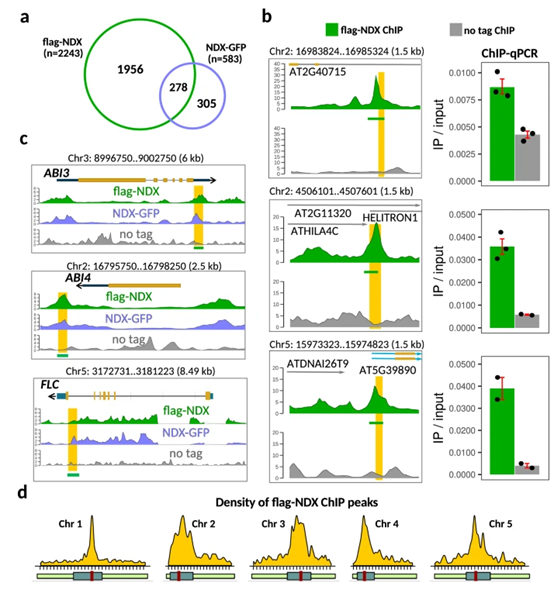
拟南芥同源异型蛋白转录因子NDX是一种核蛋白,是转录活跃的染色体臂内特定常染色体基因的调节器。本研究中作者表明NDX主要是一种异染色质调节因子,在着丝粒的周围区域起作用,来控制siRNA的产生和非CG甲基化。大多数NDX结合位点与介导转座子沉默的着丝粒周围het siRNA位点一致,并与常染色体臂中普遍存在的R-loop结构拮抗。NDX失活导致差异性siRNA积累和DNA甲基化,其中CHH/CHG低甲基化与NDX结合位点共定位。Hi-C分析显示ndx突变体的染色质结构发生了显著变化,野生型植物中ndx富集的着丝粒周围的染色体内互作信号减少,KNOT形成区域之间的染色体间互作增强,与DNA甲基化突变体中观察到的结果相似。最后得出结论,NDX是异染色质的一个关键调节因子,在功能上与het siRNA位点和非CG DNA甲基化途径耦合。