本文内容速览:
“藏粮于地、藏粮于技”是国家对农业生产领域大方向的战略方针,以科技支撑筑牢“大国粮仓”。这一战略其实也是农业领域众多科研工作者矢志不渝的奋斗方向。小远在“盘点那些与粮食作物高产相关的基因(一)”为大家总结了影响水稻产量的相关基因,“盘点那些与玉米遗传改良相关的基因”总结了影响玉米株型、产量、抗病及籽粒相关的基因,“除草剂抗性基因——作物生长的革命性转变”总结了除草剂抗性基因的研究。
老话曾说,“宁可一日无肉,不可一日无豆”。今天小远就准备围绕大豆中优良性状相关的基因展开讲述,下面就紧跟小远的步伐一起去看看吧!
GmFAD2
油酸是一种单不饱和脂肪酸,被称为“血管的清道夫”,具有降低胆固醇、血脂以及预防心血管疾病的作用。在追求健康生活的现在,高油酸花生油、菜籽油、葵花籽油、大豆油相继问世。与此同时,高油酸作物也是工业应用和生物燃料领域不可或缺的重要原料。
大豆在蛋白质食物原料和油料双重属性的加持下,成为了我国重要的农产品。高油酸大豆的开发是大豆种质改良的重要方向,大豆种子中脂肪酸生物合成途径如图1所示。大豆脂肪酸去饱和酶GmFAD2-1和GmFAD2-2是负责油酸转化的关键酶,也是创制高油酸大豆的核心关注点。美国是最早开始开展高油酸大豆种质改良的国家,可以追溯到21世纪初。
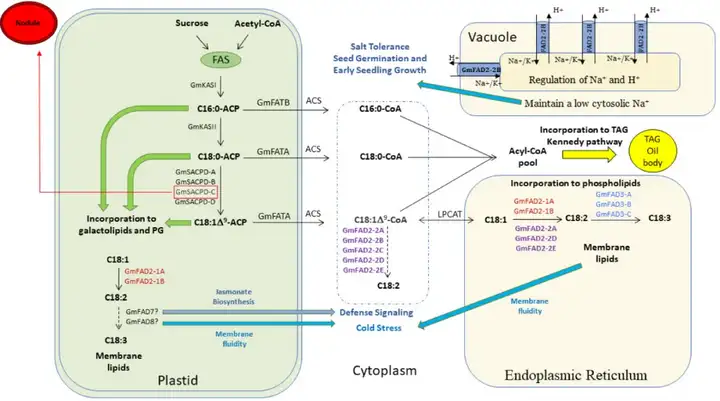
7个GmFAD2-1和GmFAD2-2成员负责将质体(GmFAD2-1A和GmFAD2-1B)、内质网(GmFAD2-1A、GmFAD2-1B、GmFAD2-2A、GmFAD2-2D和GmFAD2-2E)和细胞质(GmFAD2-2A、GmFAD2-2B、GmFAD2-2C、GmFAD2-2D和GmFAD2-2E)中的油酸(C18:0-ACP)转化为亚油酸(C18:1AΔ9-ACP)。
2008年5月,杜邦先锋公司在Food and Chemical Toxicology杂志上发表了一篇题为“Subchronic feeding study of high oleic acid soybeans (Event DP-3Ø5423-1) in Sprague-Dawley rats”的研究论文,该文中提及高油酸转基因大豆DP305423,就是围绕种子特异性ω-6脂肪酸去饱和酶的内源性GmFAD2-1基因展开的。利用RNAi技术抑制该基因的表达,从而抑制ω-6脂肪酸去饱和酶的活性,使得大豆油酸的含量得以增加。
美国明尼苏达州的Calyxt公司是一家农业生物技术公司,主要聚焦于利用基因编辑技术改善作物性状。该公司最早的成就之一就是利用TALEN技术完成了对大豆内源的两个FAD编码基因GmFAD2-1A和GmFAD2-1B的编辑,在2018年推出了首个基因编辑的商业大豆品种—FAD2KO大豆。
2020年12月,中国农科院作物科学研究所申请了一项题为“利用同时基因编辑修饰GmFAD2-1A和GmFAD2-1B获得高油酸大豆的方法”的专利。利用CRISPR/Cas9系统对受体材料中的GmFAD2-1A和GmFAD2-1B基因进行基因编辑,GmFAD2-1A和GmFAD2-1B基因均发生突变导致蛋白翻译提前终止,得到转基因植物,从而获得油酸含量达50%左右的高油酸含量的大豆突变体材料。
从上述信息中我们得知,高油酸大豆品种的创制关键在于对GmFAD2-1A和GmFAD2-1B基因的改造。因此,当需要提升大豆油酸的含量时,这些基因无疑是实现性状改良的首选目标。
GmELF3a
长童期(Long Juvenile, LJ)是指植物在达到开花潜能之前所经历的生长周期。对于大豆来说,长童期性状主要表现为在短日照条件下的晚花表型。具体来说,长童期性状的大豆在短日照条件下能够延长营养生长期,推迟开花和成熟,从而提高产量。这一性状在上世纪70年代被发现,并已成功应用于大豆的育种中。早在40多年前,遗传学研究就揭示了控制热带大豆长童期性状的J/j位点的存在,但是其背后的遗传机制当时尚未被完全解析。2016年10月,中国农业科学院作物科学研究所韩天富课题组联合华南农业大学农学院年海课题组在Molecular Plant杂志上发表了一篇题为“A single nucleotide deletion in J encoding GmELF3 confers long juvenility and is associated with adaption of tropic soybean”的研究论文,克隆了研究者们寻觅了近半个世纪的大豆长童期基因J。研究团队通过构建以“中黄24”(常规品种)和“华夏3号”(长童期品种)为亲本的重组自交系群体,采用RAD-Seq技术构建遗传图谱,进行连锁分析并结合亲本全基因组重测序,将长童期性状定位到GmELF3基因上。在长童期品种“华夏3号”中,该基因缺失了一个碱基,导致基因序列移码和所编码的蛋白失活,从而延迟开花,产生了长童期性状。将来自“中黄24”的GmELF3基因转入“华夏3号”能够实现功能互补,使“华夏3号”开花提早,从而进一步证明GmELF3突变导致长童期性状的产生。
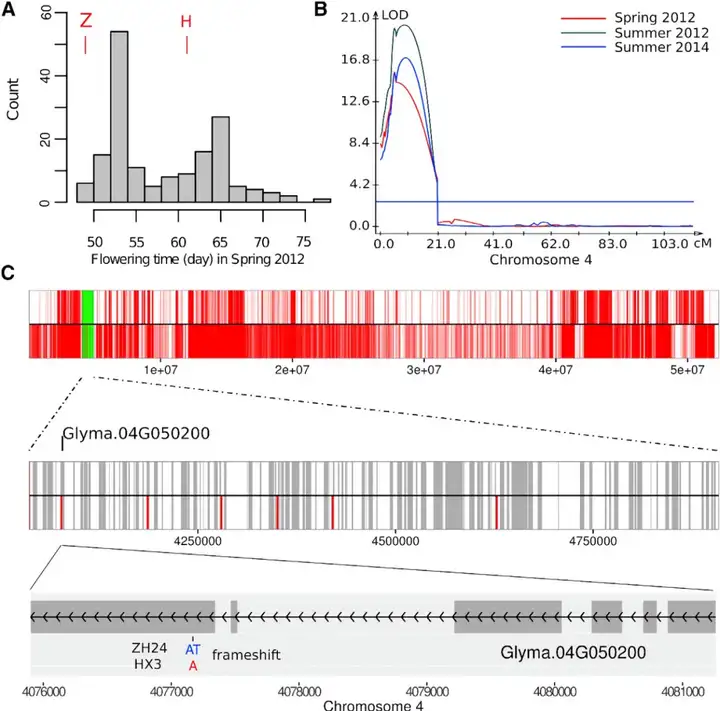
(A)中黄24×华夏3号高世代群体表现出双峰分布(2012年春季)。Z代表中黄24,H代表华夏3号;(B)4号染色体上存在一个具有表型效应的数量性状位点(QTL),该QTL是通过对中黄24×华夏3号的3个重组自交系群体使用3个季节(2012年春、2012年夏、2014年夏)的花期数据进行QTL定位而鉴定出来的;(C)亲本基因型的全基因组测序和SnpEff分析表明,只有一个多态性显著影响了QTL区域内的一个基因(Glyma.04G050200基因上的一个插入/缺失变异,位于Chr04:4077171的AT>A)。
在大豆长童期基因被初次成功克隆后,2017年3月,中国科学院东北地理与农业生态研究所孔凡江、刘宝辉课题组联合中科院遗传与发育生物学研究所田志喜课题组、中科院华南植物园侯兴亮课题组在Nature Genetics杂志上发表了一篇题为“Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield”的研究论文,该研究同样定位到了长童期性状的基因Glyma04G050200,并且揭示了大豆生态适应性相关机制。
研究团队通过正向遗传学的方法克隆了J基因(图3),发现J基因是拟南芥ELF3的同源基因,通过功能互补实验和构建近等基因系,团队对J基因在调控植物性状方面的作用进行了深入分析,包括株高、节间数、荚果数目开花时间、成熟期以及单株产量等多个表型,从而验证了J基因的功能,并且发现在低纬度条件下(短日照条件),突变型j与野生型J相比能提高大豆产量达 30-50%(图4)。
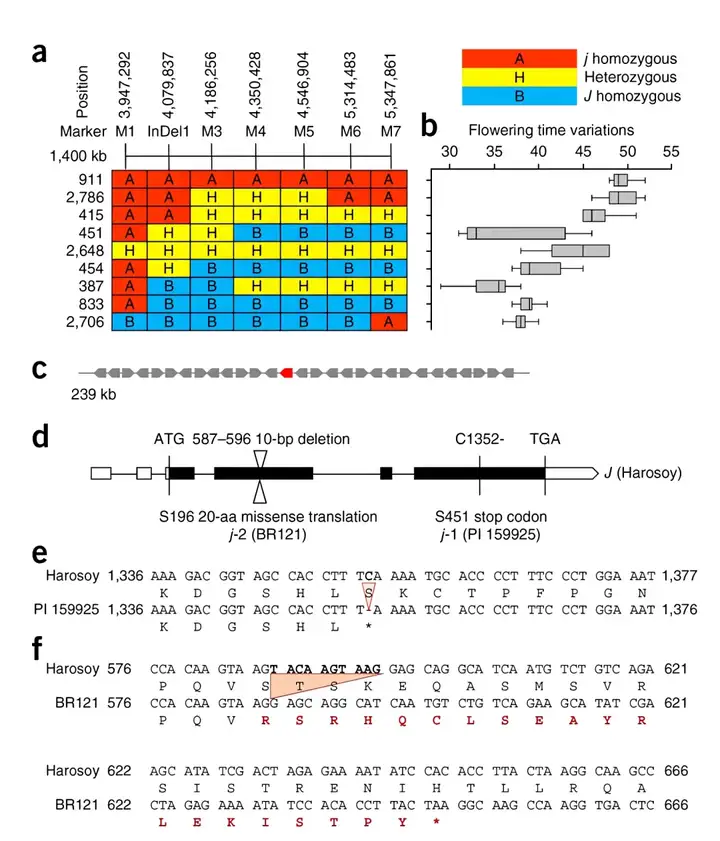
(a)BR121与Harosoy杂交的分离杂合近交群体(n=2816)中J位点在chr4 239kb区域的划分;(b)开花时间的性状分离;(c)标记M1和M3之间的239kb基因组区域包含Williams 82参考基因组中的29个预测基因;(d)J候选基因Glyma04G050200在材料PI 159925、BR121和Harosoy中的等位基因变异;(e)Harosoy与PI159925的序列比较;(f)Harosoy与BR121的序列比较。
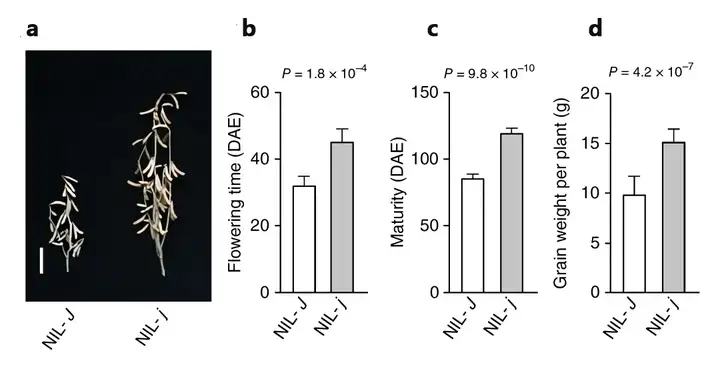
(a)NIL-J中J等位基因的存在与携带突变形式j的NIL-j相比,植株高度、节数和豆荚数减少。比例尺为10厘米;(b)开花时间;(c)成熟时间;(d)每株谷物产量。
进一步研究表明,在短日照条件下,J蛋白能够与大豆光周期开花的核心调控因子E1启动子的LUX结合元件直接结合,进而抑制E1基因的表达,从而解除了E1对FT的抑制,促进FT基因的表达上调。同时,研究还发现J基因的表达受到光敏色素蛋白E3和E4的抑制,揭示了大豆特异的光周期调控开花的遗传网络(图5)。
GmELF3a基因的克隆和作用机制的阐明不仅增进了我们对大豆生长发育调控网络的认识,而且为大豆的遗传改良和分子育种提供了新的策略和工具,这对于提高大豆的适应性、产量和品质具有深远的影响。
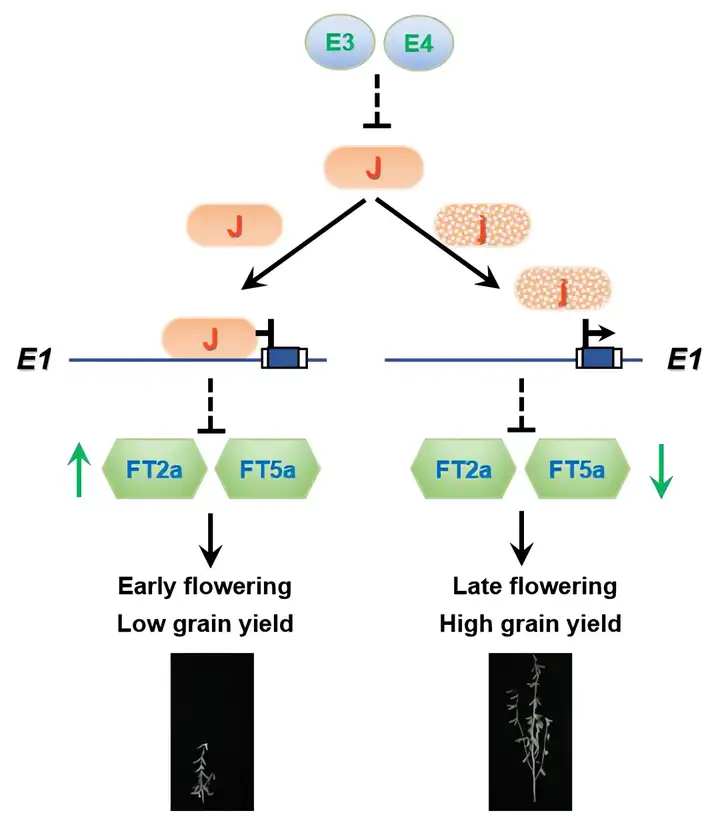
J被两个光敏色素蛋白E3和E4抑制,J蛋白与靠近LUX-binding motif的E1启动子物理结合以抑制E1转录。这减轻了E1对促进开花的FT2a和FT5a的转录抑制。当J基因功能受损时,E1不再受到抑制,继而增强了对FT2a和FT5a的抑制,导致开花时间推迟。
GmBADH1 和GmBADH2
对于不爱吃豆制品的朋友来说,豆腥味是最大的元凶。豆腥味的产生主要是由于在大豆粉碎时大豆中的脂肪氧化酶被氧气和水激活,其中的亚油酸、亚麻酸等多不饱和脂肪酸被氧化,生成过氧化物,再降解成多种具有不同程度异味的小分子醇、醛、酮、酸和胺等挥发性化合物,从而形成了大豆腥味,而且这种腥味一旦形成很难消失。
2001年,Fushimi和Masuda报道了两种本土大豆品种即Dadachamame和Chakaori,这两个品种具有令人愉悦的香味,经研究发现这是由于上述两个品种中具有2-acetyl-1-pyrroline(2AP)。2AP是一种重要的芳香化合物,在作物中能够产生如同“爆米花”的香味。其中水稻、香兰叶及面包花散发的香味也是由于该芳香化合物的存在,这一香味性状在育种中也得到了广泛的研究和应用。
2024年2月,南方科技大学朱健康课题组在Journal of Integrative Plant Biology杂志上发表了一篇题为“Breeding exceptionally fragrant soybeans for soy milk with strong aroma”的研究论文,该研究让创制香气浓郁适用于加工植物奶的专用大豆成为了可能。
研究团队以影响2AP的甜菜碱醛脱氢酶(BADH)为切入点,在已知BADH基因的变异会影响作物香气的前提下,利用基因编辑工具酶Cas12i3对大豆中BADH两个同源基因(GmBADH1和GmBADH2)进行敲除,获得了gmbadh1、gmbadh2、gmbadh1/2三种类型的功能缺失型突变体(图6A-F)。对突变体材料的表型进行了观察,结果显示其在株高、开花时间和种子重量等方面与野生型相比没有显著差异。然而,gmbadh1/2双突变体在成熟期到来的时间上明显长于野生型和其他几种突变体材料(图6G-R)。
gmbadh1/2双突植株的叶片和种子都有浓郁的稻花香味,gmbadh2单突植株稻花香味较淡,gmbad1单突和野生型中都闻不到稻花香味。分析显示,gmbadh1/2双突变的叶片和种子中,2AP含量大幅增加,是gmbadh2单突变体的4倍多(图6S、T),而在gmbad1单突和野生型中都检测不到2AP。这表明两个GmBADH基因功能冗余,GmBADH2在控制大豆中2AP含量方面发挥着主要作用,GmBADH1对这种控制也有显著贡献。由gmbadh1/2双突变体大豆制成的豆乳表现出更强的香味,这与2AP水平一致(图6U)。参与研究的志愿者在选择不同原料制成的豆乳时,会更偏好选择香气增强的gmbadh1/2大豆制成的豆乳(图6V)。
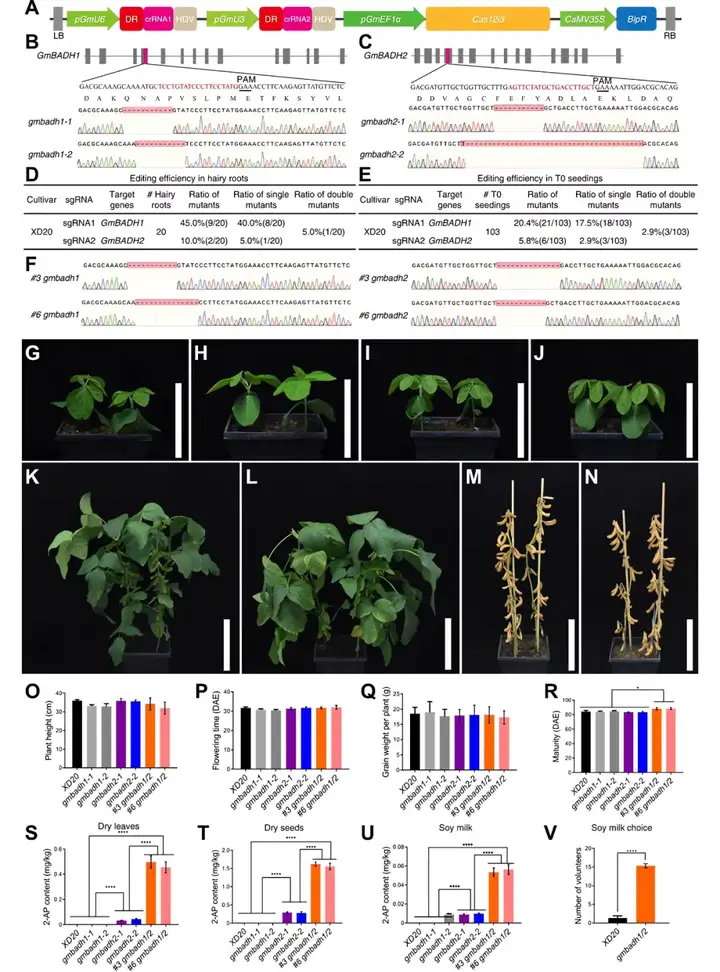
(A)CRISPR/Cas12i3构建示意图;(B)GmBADH1 crRNA设计和突变基因型;(C)GmBADH2 crRNA设计和突变基因型;(D、E)毛状根转化系统和基因编辑系统的编辑效率;(F)双突变基因型;(G、K、M)野生型XD20分别在16DAE(days after emergence)、74DAE及98DAE时的表型;(H)gmbadh1-1(左)和gmbadh1-2(右)在16DAE时的表型;(I)gmbadh2-1(左)和gmbadh2-2(右)在16DAE时的表型;(J、L、N)gmbadh1/2双突变体#3(左)和#6(右)分别在16DAE、74DAE及98DAE时的表型;(O-R)野生型XD20和突变体的株高(O)、开花时间(P)、种子重(Q)和成熟期(R);(S-U)野生型XD20及其突变体干叶(S)、干种子(T)和豆浆(U)中2-AP含量;(V)选择野生型XD20或gmbadh1/2双突变体豆浆的志愿者人数。
这篇文章向我们展示了香味大豆实现的可行性策略,创制香味大豆不仅能够提升豆制食品的风味和营养价值,还能够促进农业和食品工业的发展,增加农民收入,满足消费者对高品质食品的需求。
GmRIC1/2 和POWR1
大豆是重要的植物蛋白来源,大豆蛋白包含8种人体必需的氨基酸,属于优质蛋白,可以作为肉类的良好替代品。对于素食主义者或者健身人士是一种很好的选择。高蛋白育种是大豆育种的关键领域,然而大豆蛋白质的遗传改良进程相对缓慢。这一现象主要是由于大豆蛋白质与油脂之间存在极显著的负相关性,即在提升大豆蛋白含量的过程中,油脂含量往往会相应减少。鉴于此,众多科研工作者仍在不懈努力,致力于培育出既富含蛋白质又保持适宜油脂含量的大豆新品种。
2022年4月,中国科学院东北地理与农业生态研究所张恒友课题组在Nature Communications杂志上发表了一篇题为“POWR1 is a domestication gene pleiotropically regulating seed quality and yield in soybean”的研究论文,该研究进行了大豆高蛋白主要控制基因的鉴定和分析。
研究团队利用全基因组测序在20号染色体上鉴定到了一个主效QTL-cqPro-20,并发现该QTL的候选基因是一个编码CCT结构域的基因POWR1(Protein, Oil, Weight Regulator 1),其中321bp转座子在CCT结构域的插入/缺失是该位点引起蛋白和油脂变异的主因(图7)。
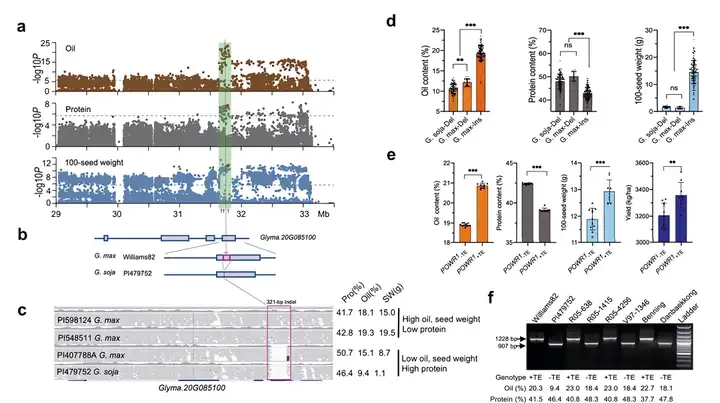
(a)曼哈顿图显示了油脂、蛋白质含量和百粒重与chr20上4Mb区间内SNPs和InDels的关系;(b)Glyma.20G085100含和不含321bp InDel的基因结构(红框);(c)Glyma.20G085100的两个高油/低蛋白序列和两个低油/高蛋白序列的测序结果显示其321bp的InDel;(d、e)321bp插入/缺失(InDel)等位效应在关联小组中对油脂、蛋白质、百粒重和产量的影响(对于大豆G. soja-Del、G. max-Del和G. max-Ins分别为116、8和154个样本)以及在近等基因系(NILs)中的影响(n=12);(f)4个RIL群体亲本转座元件插入的PCR基因分型分析。
让我们把视角转向另一个研究团队,大豆高蛋白育种比较困难,除蛋白与油脂有显著的负相关性外,蛋白与产量通常也会存在这样的负相关性。该团队另辟蹊径,以优化结瘤固氮促进高产优质为思路展开了研究。
2024年5月,广州大学关跃峰课题组、孔凡江课题组联合中科院分子植物科学卓越创新中心王二涛课题组在Nature Plants上发表了一篇题为“Genetically optimizing soybean nodulation improves yield and protein content”的研究论文,该研究通过基因编辑精准调控根瘤数量,实现碳氮平衡的高效固氮,从而在大田种植条件下大幅提高大豆产量和蛋白含量。
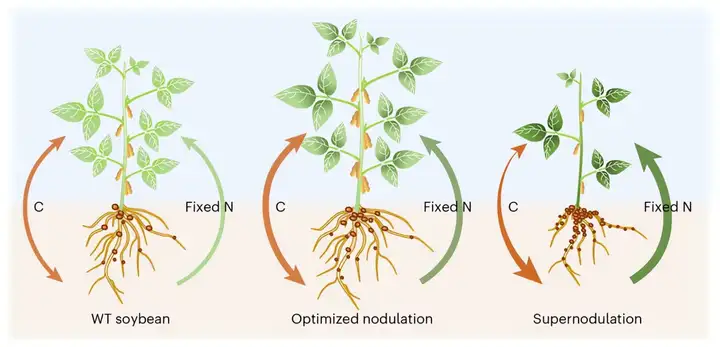
在“大豆多结瘤会减产”的思维定势下,该团队创造性地提出了一个假设:在优化根瘤数量的情况下,根瘤中氮获取能力的提高和碳消耗的增加可能会达到平衡,从而仍然有利于茎的生长。为了验证该假设,研究团队利用基因编辑创造了根瘤数量不同程度改变的各种大豆突变体(nin-4m,ric1b/2b,ric1a/2a,ric-6m和nark),发现超级结瘤大豆突变体ric-6m和nark生物量减少,而根瘤增加1倍的ric1a/2a突变体生物量显著增加(图8a-c)。同位素示踪等实验表明,ric1a/2a根瘤数量适当提高,不仅增加生物固氮作用,还通过氮素增加了叶绿素含量,增强大豆光合效率,最终达到碳氮协同促进(图8d-i)。
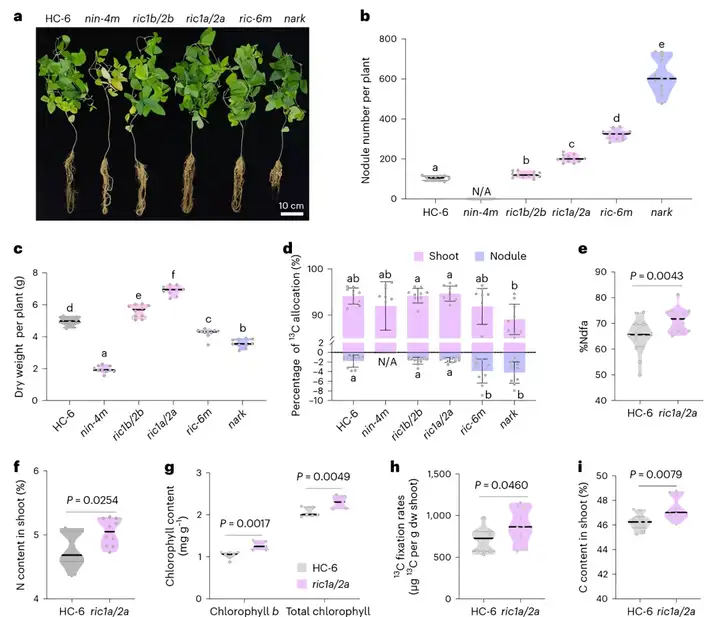
(a-c)接种40d后,ni-4m、ric1b/2b、ric1a/2a-1、ric1a-6m、ark和HC-6水培植株形态(a)、根瘤数(b)和全株干重(c)(n=12);(d)单株根茎组织中总13C(n=10);(e)来自大气的氮(%Ndfa)的百分比,代表ric1a/2a-1和HC-6(n = 15)植株中根瘤固氮的贡献;(f)HC-6和ric1a/2a突变体植株茎部组织总氮含量(n=10);(g)HC-6和ric1a/2a突变体植株茎部组织叶绿素含量(n=10);(h)HC-6和ric1a/2a突变体植株茎部组织13C固定率(每克干重(dw)茎部每12h)(n=10);(i)HC-6和ric1a/2a突变体植株茎部组织总C含量(n=10)
为了测试ric1a/2a的农艺性状,研究团队也进行了多年多点的田间试验(图9a-d)。与野生型HC-6相比,ric1a/2a的小区产量显著提升10%-20%以上,蛋白质含量稳定提高1-2个百分点,且不显著降低含油量(图9e、f)。
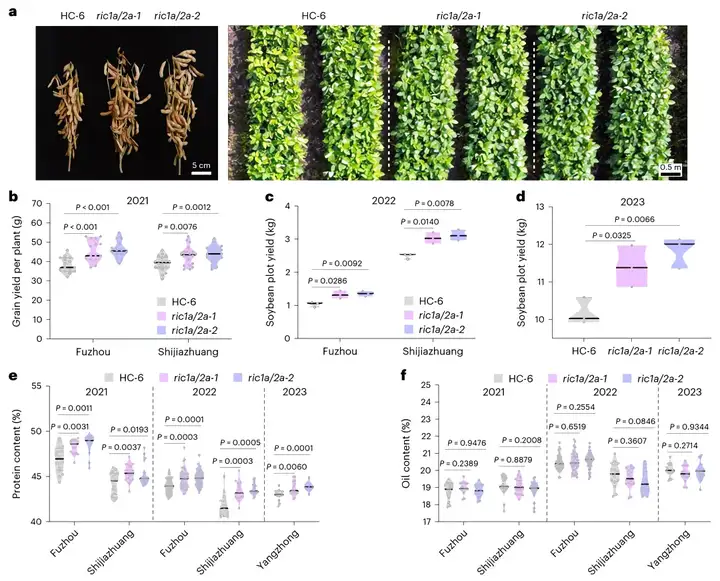
(a)左图为2023年在洋中种植的成熟HC-6和两株ric1a/2a突变株的表型。右图是无人机对田间种植植物的鸟瞰图。左边两行为HC-6;右边的四行分别是ric1a/2a-1和ric1a/2a-2;(b)2021年福州和石家庄种植的HC-6和ric1a/2a突变体单株产量;(c)2022年福州(6.75 m2)和石家庄(7 m2)HC-6和ric1a/2a小区产量(n=3);(d)2023年洋中地区HC-6和ric1a/2a突变体亩产(50 m2)(n=3);(e、f)在所有试验中,大豆蛋白含量(e)和油脂含量(f)的数据如下:2021年福州试验点16个样本,2021年石家庄试验点18个样本,2022年福州试验点40个样本,2022年石家庄试验点16个样本,2023年洋中试验点15个样本。
研究分析表明,出现这种现象的原因可能在于该突变体与那些会导致大量碳消耗、生长发育受阻和产量降低的超结瘤大豆突变体不同。通过基因编辑技术对大豆结瘤过程进行优化,该突变体成功维持了碳氮的平衡分配,加强了碳氮同化的效率,从而促进了茎部的生长,改善了整体的生长发育状况。
『小远叨叨』
在这篇文章中,小远为大家精心梳理了大豆育种领域中备受关注的性状改良需求,以及与之关联的关键基因。文章还揭示了基因编辑技术在实现这些性状改良中的潜力。农业农村部发布的两项转基因生物安全证书,也向育种专家们传递着积极的信号。小远在与公司基因编辑研发负责人的深入交流中,听到了一句令人印象深刻的话:“在求学时期,我们一直强调产学研结合的重要性;现在步入职场,我更加渴望能将所学知识转化为实际应用。”我们拥有一群有志向、有能力的团队成员,一个值得信赖的公司平台,以及坚实的技术基础。我们热忱欢迎有合作意向的专家学者前来洽谈,共同推进育种事业的发展!

References:
Lakhssassi N, Lopes-Caitar VS, Knizia D, et al. TILLING-by-Sequencing+ Reveals the Role of Novel Fatty Acid Desaturases (GmFAD2-2s) in Increasing Soybean Seed Oleic Acid Content. Cells. 2021;10(5):1245.
Delaney B, Appenzeller LM, Munley SM, et al. Subchronic feeding study of high oleic acid soybeans (Event DP-3Ø5423-1) in Sprague-Dawley rats. Food Chem Toxicol. 2008;46(12):3808-3817.
Yue Y, Liu N, Jiang B, et al. A Single Nucleotide Deletion in J Encoding GmELF3 Confers Long Juvenility and Is Associated with Adaption of Tropic Soybean. Mol Plant. 2017;10(4):656-658.
Lu, S., Zhao, X., Hu, Y. et al. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat Genet. 2017;49, 773–779.
Fushimi T, Masuda R. 2-Acetyl-1-pyrroline concen-tration of the vegetable soybean. In: Lumpkin T, Shanmugasundaram S (eds) Proceedings of the 2nd international vegetable soybean conference. Washington State University, Pullman, Washington, USA, p 39. 2001.
Xie H, Song M, Cao X, et al. Breeding exceptionally fragrant soybeans for soy milk with strong aroma. J Integr Plant Biol. 2024;66(4):642-644.
Goettel W, Zhang H, Li Y. et al. POWR1 is a domestication gene pleiotropically regulating seed quality and yield in soybean. Nat Commun. 2022;13, 3051.
Zhong X, Wang J, Shi X. et al. Genetically optimizing soybean nodulation improves yield and protein content. Nat. Plants. 2024, 10, 736–742.
