本文内容速览:
提高作物抗病性一直是育种学家研究的热点,同时也是确保作物高产和品质安全的重要指标。实际的农业生产表明培育高抗病性的作物品种是防御病虫害最经济有效的途径,通过各种生物学技术,利用广谱抗性(BSR)的抗性基因(R基因)可培育出商业化的抗病品种。然而鱼和熊掌不可兼得,植物自身的资源是有限的,引入R基因获得高抗病性的同时,往往会伴随着其他优良性状的丧失;反之植物快速生长往往伴随着对病原物易感性的增加、非生物胁迫耐受性的降低,这种现象称为植物生长-防御平衡。植物只有将生长和防御二者平衡才能在自然界中更好地生存和繁殖,因此植物生长-防御平衡的实现是创制同时具有高产和高防御能力作物的有效策略。接下来,跟着小远一起看看与植物生长-防御平衡相关的基因和育种策略吧!
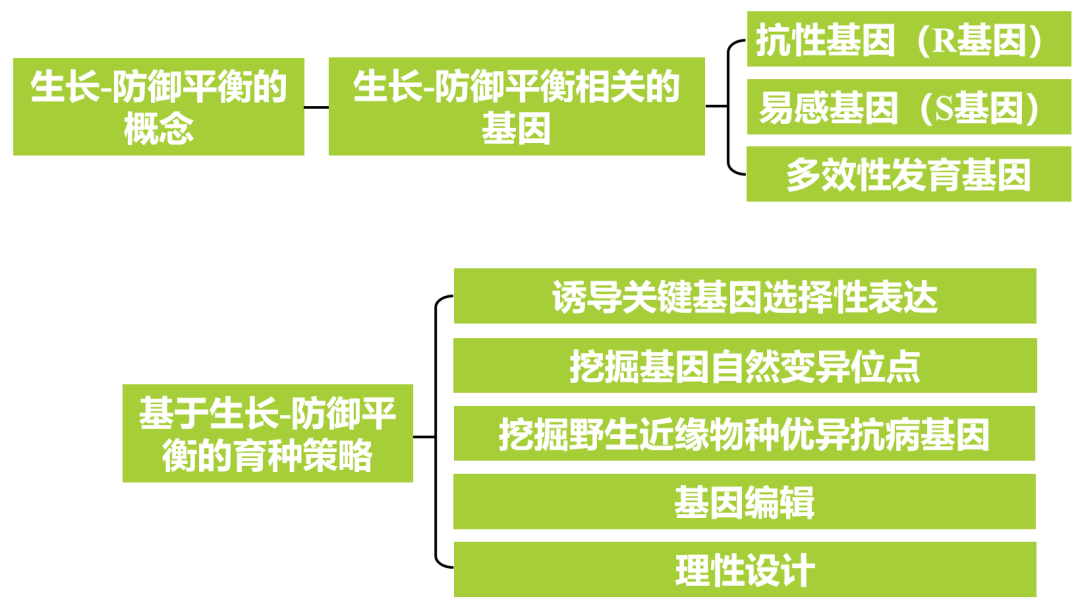
2017年,中国科学院上海植物生理生态研究所何祖华课题组在Science杂志上发表了题为“Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance”的研究论文。在该研究论文中,作者发现了水稻Pigm抗性位点包含一个编码核苷酸结合的NLR受体的基因簇,该受体赋予水稻对稻瘟病菌的持久抗性而不影响产量。在该簇中,PigmR赋予水稻BSR,其在所有组织中低水平表达并降低籽粒重量;PigmS是一个感病基因,在花粉中高度表达,可以增加种子结实(图1A、B,图2)。PigmS启动子含有两个串联微型转座子MITE1和MITE2,其触发RNA介导的DNA甲基化,导致叶子中的PigmS表达,从而导致植株感病。PigmS与PigmR相互作用从而破坏PigmR的功能,实现水稻生长和抗病性之间的平衡,即生长-防御平衡。
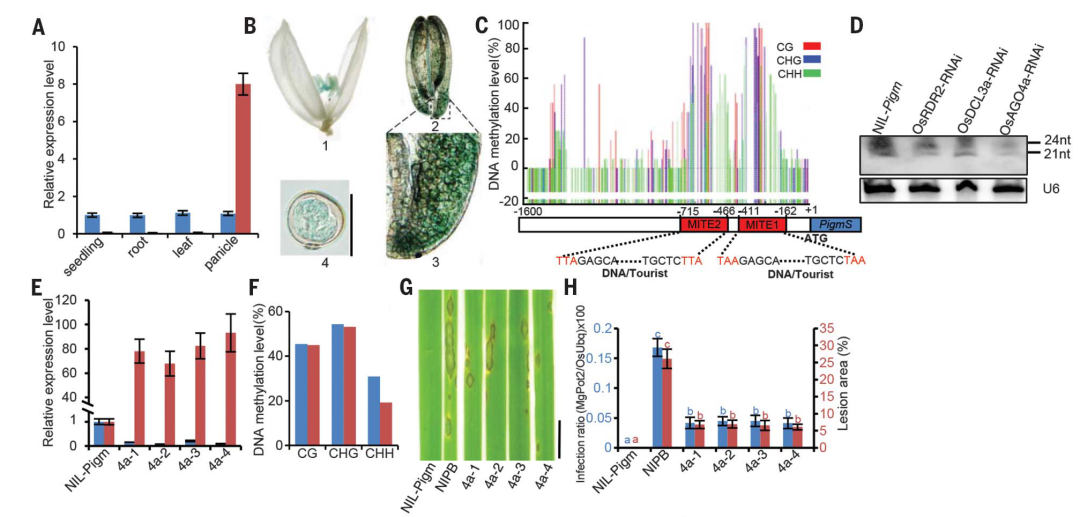
图1 PigmS的表达受RNA介导的DNA甲基化(RdDM)途径调控(Deng et al., 2017)。(A)以水稻OsActin1为对照,通过qRT-PCR检测PigmR(蓝色)和PigmS(红色)的表达;(B)在pPigmS::GUS花药中检测到GUS活性,表明花粉中PigmS高表达,1:小穗、2:花药、3:花药的一部分、4:GUS染色花粉;(C)叶片中PigmS启动子的CG、CHG和CHH甲基化及PigmS启动子MITE1和MITE2的示意图;(D)与野生型NIL-Pigm相比,RdDM途径RNAi突变体中MITE1和MITE2衍生的24nt的siRNA的积累减少,U6作为对照;(E)qRT-PCR检测4个OsAGO4a-RNAi细胞系中OsAGO4a(蓝色)和PigmS(红色)的转录本水平;(F)与野生型系(蓝色)相比,OsAGO4a-RNAi叶片中PigmS启动子CHH甲基化降低(红色);(G、H)OsAGO4a-RNAi株系感病性降低。
注:RdDM和RNAi途径均产生siRNA,并涉及保守的AGO、Dicer和RNA依赖的RNA聚合酶蛋白(Erdmann R M and Picard C L,2020)。
图2 PigmS抵消了PigmR介导的高抗病性而导致的产量损失(Deng et al., 2017)。(A)非病害条件下田间试验中转基因植物的产量组成。与野生型日本晴(NIPB)相比,PigmR株系的千粒重(红色)显著降低,而PigmS、PigmR和PigmS双转基因株系的结实率(绿色)显著提高。由此产生的单株重量(蓝色)在PigmR和PigmS品系中分别显著降低和提高。PigmR和PigmS同时发挥作用不影响粮食产量;(B)在自然非病害条件下(绿色)和在稻瘟病侵染条件下(红色),NILs的实际产量。
综上,我们可以知道R基因通常是成对发生作用,在没有病原物的情况下防止自身免疫和减少防御成本,在病原体感染时实现有效的免疫反应来介导生长-防御的平衡。这里只给大家介绍了一种DNA甲基化介导的R基因的微调,其他例如蛋白修饰等也能实现R基因的微调,这里主要是为了告诉大家可以通过微调R基因从而实现生长-防御平衡,因此对于如何利用R基因实现生长-防御平衡的方法就未过多介绍,感兴趣的小伙伴可以自己查阅相关文献。
2021年中国科学院植物生理与生态研究所何祖华课题组在Cell杂志上发表了一篇题为“Ca2+sensor-mediated ROS scavenging suppresses rice immunity and is exploited by a fungal effector”的研究论文,作者发现ROD1是一种新型的钙离子传感器,在水稻中编辑ROD1基因提高了水稻对稻瘟病、纹枯病等多种病害的BSR。
2022年,中国农业科学院植物保护研究所宁约瑟课题组在Cell Reports杂志上发表了一篇题为“A VQ-motifcontaining protein fine-tunes rice immunity and growth by a hierarchical regulatory mechanism”的研究论文,作者发现OsVQ25可被OsPUB73泛素化降解,可通过抑制OsWRKY75的转录活性从而负调控水稻的BSR,敲除OsVQ25可提高水稻对稻瘟病菌和细菌性枯萎病的抗性,但不影响水稻的生长。
2022年西北农林科技大学王晓杰课题组在Cell杂志上发表了一篇题为“Inactivation of a wheat protein kinase gene confers broad-spectrum resistance to rust fungi”的研究论文,作者发现小麦PslPK1基因可磷酸化免疫调节转录因子CBF1d,并抑制其功能,敲除PslPK1可提高小麦对锈病的抗性,但不影响小麦的生长。
2023年,华中农业大学李国田课题组在Nature杂志上发表了一篇题为“Genome editing of a rice CDP-DAG synthase confers multipathogen resistance”的研究论文,作者发现RBL1参与植物中磷脂的生物合成,通过基因编辑敲除RBL1中的12个碱基对可以产生一个优异的变异等位基因,该等位基因在赋予BSR的同时不会影响作物产量。
综上,对S基因的编辑是实现植物生长-防御平衡的重要途径,也是创制具有高抗病性、高产量品种的重要方法。
1.3 多效性发育基因
多效性发育基因指的是在植物生长发育的过程中可以影响多个生物学过程,进而影响多个性状的基因。
IPA1是一个典型的多效性发育基因,相关的研究可以查看“转录因子IPA1,次次都能发顶刊是怎么做到的?”在水稻中,ipa1-1D和ipa1-2D等位基因可提高IPA1的表达,从而提高产量与籽粒重量(图3),同时IPA1还可以激活免疫相关基因WRKY45的表达来提高抗病性,从而保持生长和防御之间的平衡(Wang et al., 2018,Zhang et al., 2017)。除IPA1外,由Gα、Gβ和Gγ亚基组成的异源三聚体G蛋白,已经在玉米中被证明可以同时调节枝条分生组织发育和免疫反应(Wu et al., 2020)。
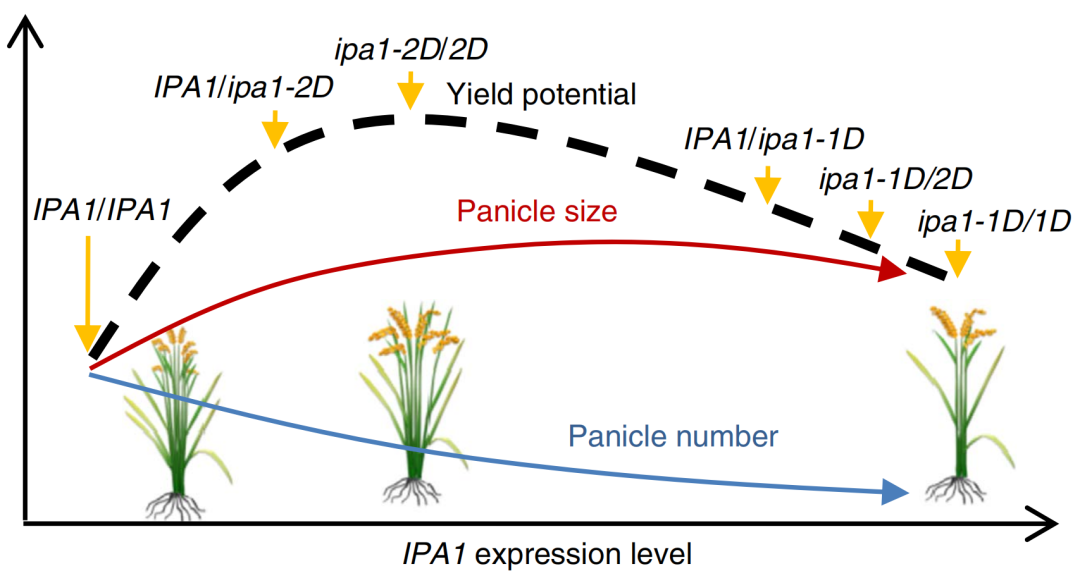
图3 IPA1、ipa1-2D和ipa1-1D等位基因组合形成的产量潜力模型(Zhang et al., 2017)。该模型预测,IPA1表达量适中、穗数较大、分蘖数适中的植株产量较高。
MicroRNA是植物中各种性状的关键调节因子,它可以影响植物的多种表型性状并且保持生长-防御平衡(Tang et al., 2021)。2021年,四川农业大学王文明课题组在Nature Plants杂志上发表了一篇题为“Suppression of rice miR168 improves yield, flowering time and immunity”的研究论文,作者发现miR168靶向编码RNA诱导沉默复合物的关键成分AGO1,敲除miR168可以增加稻穗数量、减少开花时间从而提高水稻产量同时提高水稻的抗病性。实验结果表明通过关键的miRNA可以同时控制多个性状,维持植物的生长-防御平衡。
以上,小远简要的介绍了与生长-防御平衡相关的三种类型的基因,并列举了相关的文献案列,感兴趣的小伙伴可以下载阅读学习其中的研究思路。接下来一起看看基于生长-防御平衡的育种策略吧!
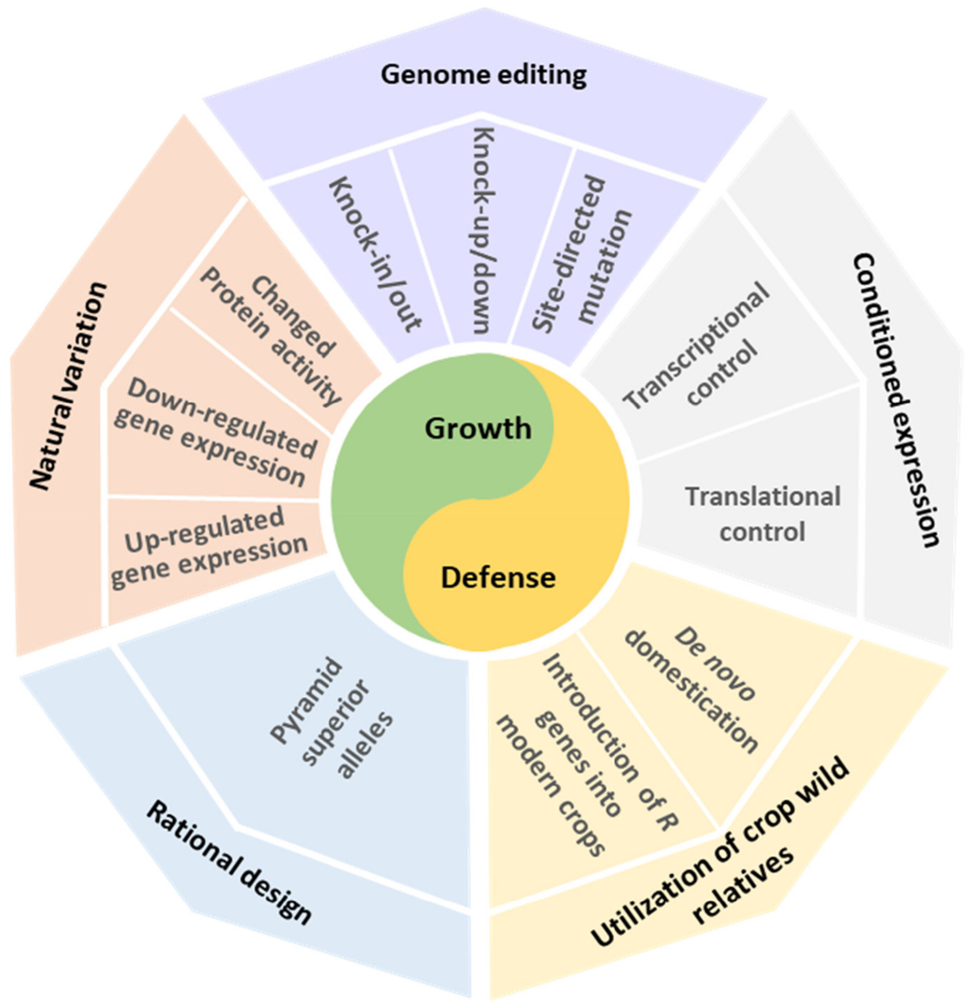
传统的育种策略为植物抵抗病害提供了有效的解决方法,然而传统的育种策略一般是费时费力的。随着生物技术的发展,多种新兴的育种技术被提出,感兴趣的小伙伴可以查看“青出于蓝-单倍体诱导技术的衍生”、“纯系养成—单倍体诱导技术(一)”、“纯系养成—单倍体育种技术(二)”。多个R基因、S基因及多效性发育基因也已经被鉴定出来,并且相关的功能也已经被证明。然而,由于植物生长-防御平衡的存在,这些基因需要通过恰当的分子生物学技术进行微调。2024年,中国科学院植物生理生态研究所何组华课题组在Plant Biotechnol Journal杂志上发表了一篇题为“Revisiting growth-defence trade-offs and breeding strategies in crops”的研究论文,作者提出了基于生长-防御平衡的五种潜在的育种策略(图4)。
此外,翻译调控也是诱导基因选择性表达的有效方法。基于翻译起始模型,到达下游的mORF起始密码子只能通过漏扫描uORF的ATG或翻译后重新启动(Yuan et al., 2023)。因此终止密码子位于mORF之前的Type1-uORF可通过漏扫描和再起始进行mORF翻译;终止密码子位于mORF起始密码子之后的Type2-uORF只能通过漏扫描进行mORF的翻译(图5A)。理论上,没有uORF(ORF-free)、有Type1或Type2 uORF的5′ UTR会产生不同程度的翻译抑制。2024年,武汉大学胥国勇课题组和华中农业大学袁猛课题组合作在Science China Life Sciences杂志上发表了一篇题为“Engineering disease-resistant plants with alternative translation efficiency by switching uORF types through CRISPR”的研究论文。在该研究论文中,作者首先克隆了拟南芥TBF1的5′ UTR的uORF并且通过改变uORF的类型结合选择性核糖体印迹技术发现uORF-free、Type1/2-uORF对TBF1的翻译效率具有不同的抑制作用(图5A-D)。作者根据此实验结果开发出了uORF转换系统(CRISPR-aTrE-uORF)(图5E),并利用该方法在水稻中筛选到了15个抗病相关基因,4个感病相关基因(图5F)。随后作者选取了谷氨酰胺合成酶2(OsGS2)基因进行了抗病性和生长的检测,发现将其Type1-uORF切换为Type2-uORF时,提高了对GS2的翻译抑制,从而在不影响生长的前提下提高了水稻的抗病性(图6、7)。该文献的研究结果表明CRISPR-aTrE-uORF可以同时实现未知抗病相关基因的挖掘和表达水平的有效控制,并有望应用于玉米、小麦、大豆等其他作物中。
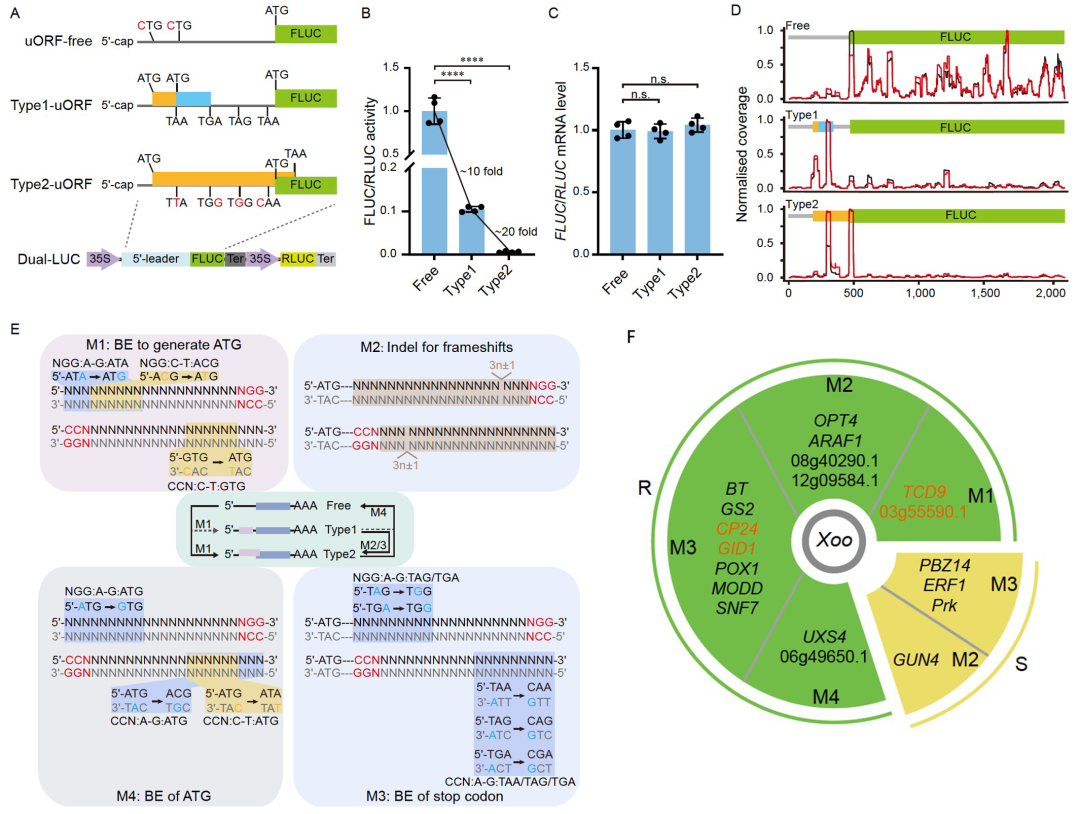
图5 改变uORF类型的方法(Tian et al., 2024)。(A)携带不同uORF类型的5′ UTR介导的FLUC活性的翻译控制;(B、C)FLUC/RLUC活性和mRNA水平;(D)选择性核糖体序列显示核糖体在uORF和FLUC上的结合模式,红线:生物素标记靶探针富集的选择性核糖序列,黑线:未富集的Ribo-seq;(E)用CRISPR切换uORF类型的方法。M1:通过A-G或C-T碱基编辑方法生成ATG,M2:indel碱基通过CRISPR介导indel方法在Type1-uORF内引入3n±1个移码;M3:A-G碱基编辑法去除Type1-uORF终止密码子;M4,通过A-G或C-T碱基编辑方法删除uORF的ATG;(F)用M1-M4方法鉴定出15个抗性基因(R:绿色背景)和4个易感基因(S:黄色背景)。具有明显生长或发育缺陷的aTrE等位基因为橙色。
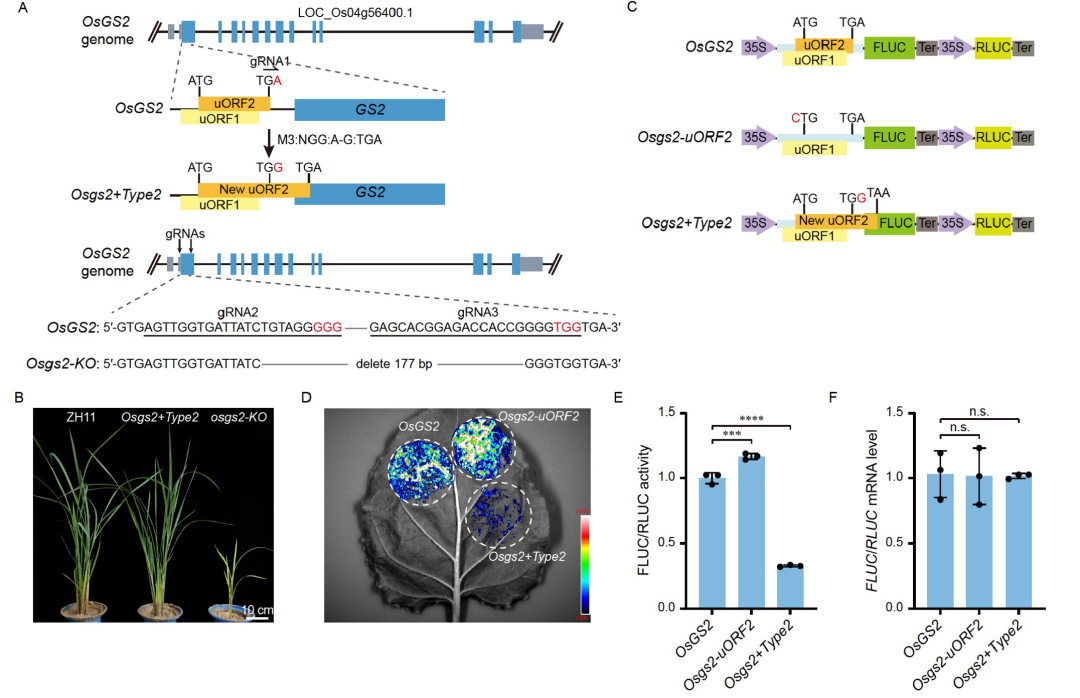
图6 Osgs2+Type2–uORF翻译效率检测(Tian et al., 2024)。(A)Osgs2+Type2–uORF和Osgs2–KO突变体示意图;(B)Osgs2+Type2–uORF和Osgs2–KO突变体的整体生长;(C)双荧光素酶系统测量WT、Osgs2+Type2–uORF和Osgs2–uORF2的5′ UTR翻译效率示意图;(D-F)LUC活性的拍照示意图(D)、FLUC/RLUC活性(E)和瞬时表达后的mRNA水平(F)。
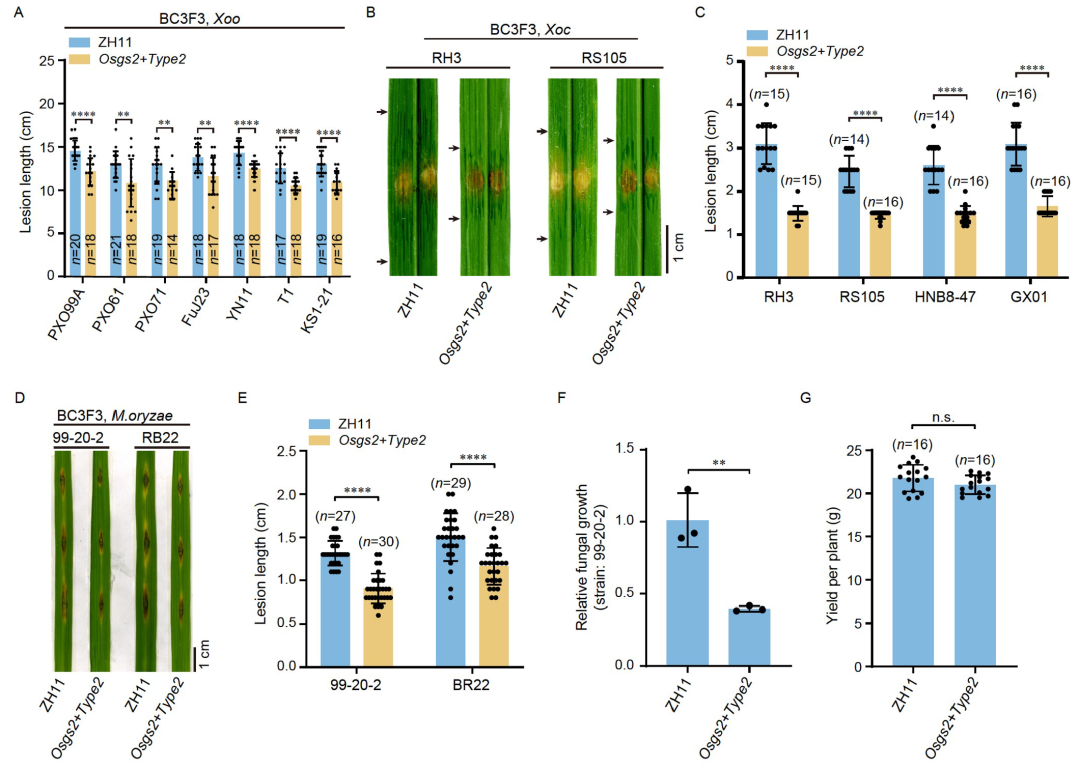
图7 Osgs2+Type2–uORF的广谱抗病性检测(Tian et al., 2024)。(A)细菌病原体黄单胞菌(Xoo)感染Osgs2+Type2基因的定量分析。菲律宾PXO99A、PXO61、PXO71,日本T1,中国FuJ23和YN1这6个水稻品种为实验组,ZH11为对照;(B、C)水稻细菌性条斑病(Xoc)在生长室内感染后的症状(B,水浸泡)和定量(C)。RH3、RS105、HNB8-47和GX01为不同的Xoc菌株;(D-F)稻瘟病在生长室内感染后的症状和定量。99-20-2和BR2代表不同的稻瘟病菌类型,对病变长度(E)和真菌量(F)进行量化;(G)田间试验的单株产量。
注:BC3F3代表Osgs2+Type2–uORF与亲本ZH11回交3代后自交3代。
策略二:挖掘基因自然变异位
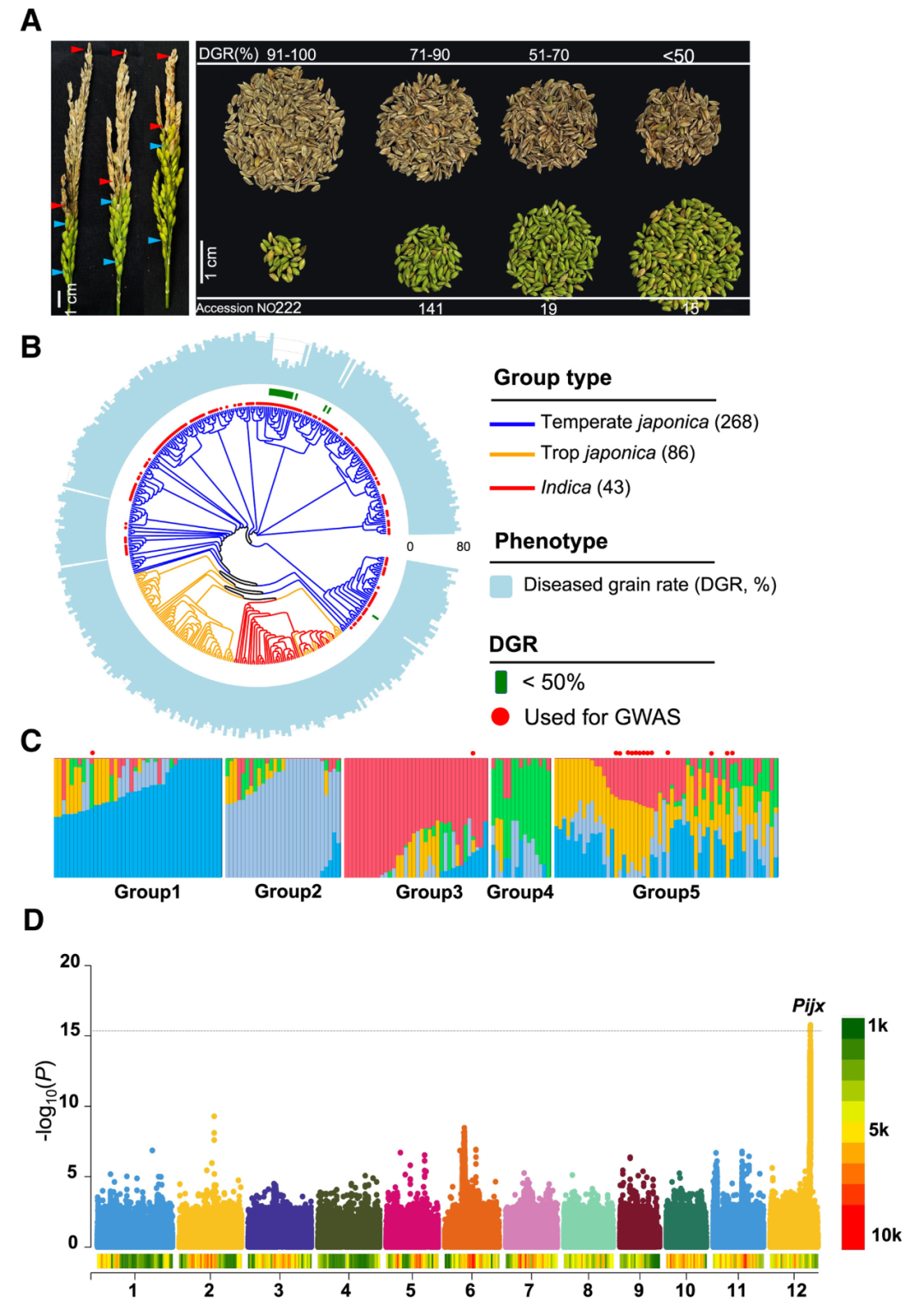
物竞天择,适者生存。优良的自然变异会经过自然适应和人工选择,自然变异具有抑制基因表达、激活基因表达、改变蛋白质活性等功能。从生长-防御平衡的角度来看,探索基因的自然变异位点是宝贵的育种资源。2022年,中国农业大学李自超课题组在Nature Communications杂志上发表了一篇题为“Natural variation of DROT1 confers drought adaptation in upland rice”的研究论文,作者首先利用全基因组关联分析(GWAS)鉴定了一个与水稻耐旱性相关的关键基因DROT1并验证其确实可提高水稻耐旱性。随后为了研究DROT1在种质资源中的自然变异并鉴定优良等位基因,作者对743个水稻样品进行了单倍型分析,一共发现了8个单倍型,其中Hap3在旱稻中的占比远高于其他单倍型,DROT1Hap3启动子区的关键SNP由C突变为T提高了DROT1在旱稻中的表达量,从而提高了水稻的耐旱性(图8)。
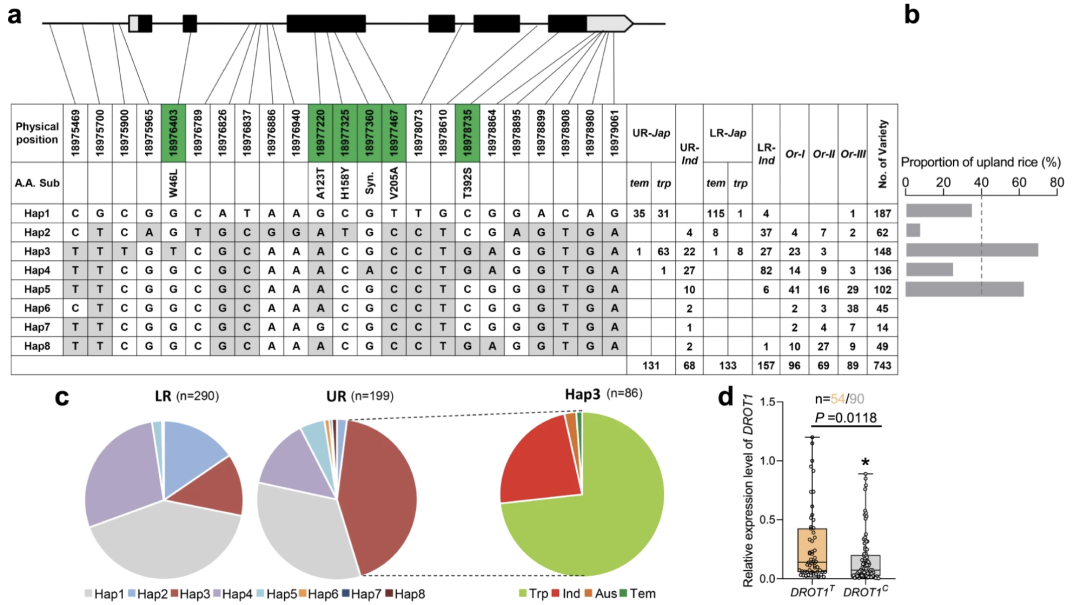
图8 DROT1的单倍型分析(Sun et al., 2022)。(a)自然群体中DROT1的单倍型。编码区的核苷酸变异用绿框标记,每个单倍型(Hap1-8)品种数量显示在右栏中。UR:旱地水稻,LR:低地水稻;(b)具有Hap1-Hap5基因的旱地水稻品种比例。虚线表示旱地水稻在整个品种种群中的平均比例;(c)DROT1单倍型在低地水稻和旱地水稻中的分布;(d)干旱胁迫下DROT1在DROT1T型和DROT1C型植株中的相对表达量。DROT1C为非Hap3单倍型。
策略三:挖掘野生近缘物种优异抗性基因
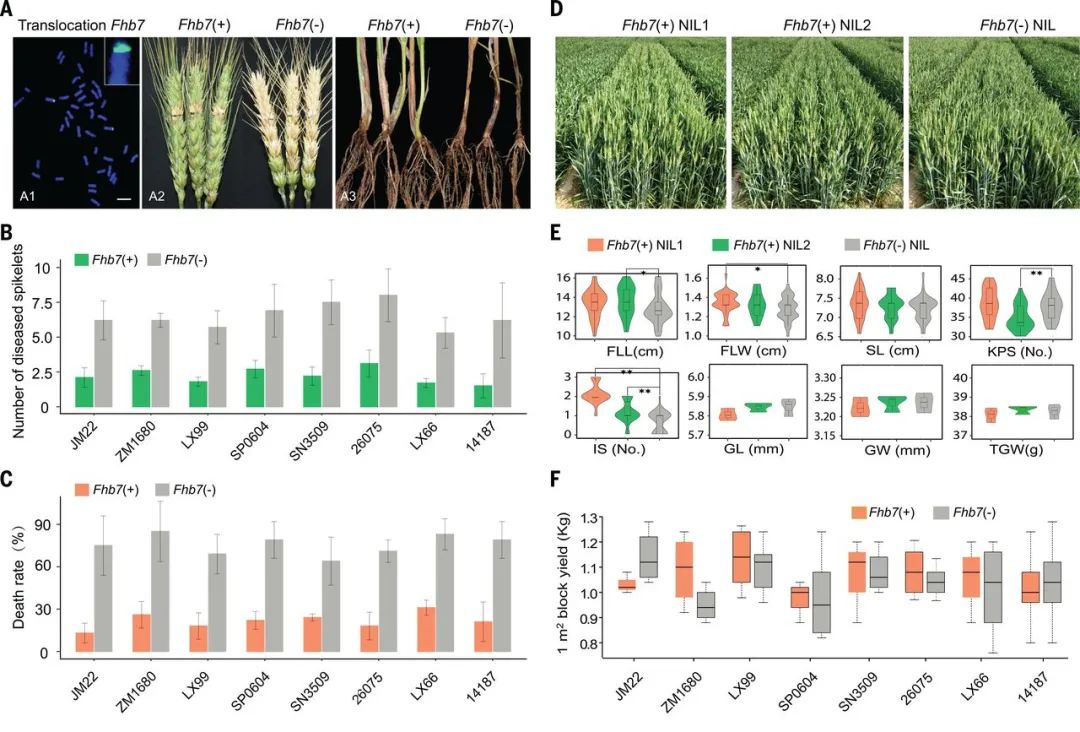
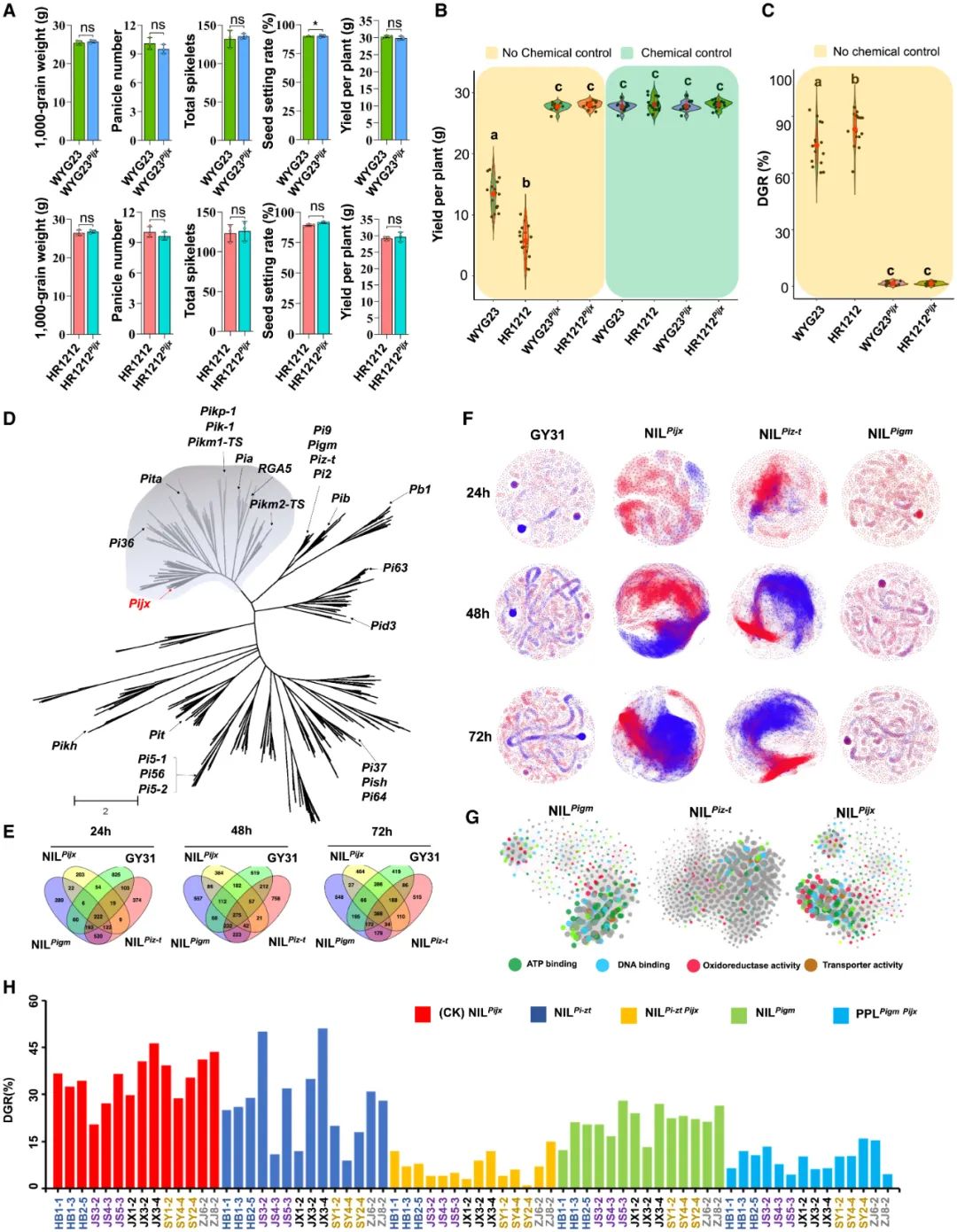
作物野生近缘物种(CWRs)的遗传多样性远高于人工选育种质的遗传多样性,因此CWRs是作物遗传改良宝贵的基因资源库。最直接利用CWRs的方法是将从CWRs种挖掘出来的优良R基因引入栽培物种中。2020年,山东农业大学孔令让课题组在Science杂志上发表了一篇题为“Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat”的研究论文,作者从小麦野生近缘种Thinopyrum elongatum的基因组中克隆了一个编码谷胱甘肽S-转移酶的Fhb7基因,并将其转入栽培品种小麦中,发现Fhb7可以提高小麦对赤霉病的抗性,但不会影响小麦产量(图9)。
策略四:基因编辑
基因编辑技术可以实现基因的突变、缺失、敲入、敲高或关键位点的突变,并且近年来已经逐渐应用于育种中。利用基因编辑技术可以有效的编辑S基因,从而创制具有生长-防御平衡的作物品种(Yu et al., 2021)。编辑S基因相关的文献,在本文的上一部分已经给大家列举了一些,这里就不再进行例举。
References:
