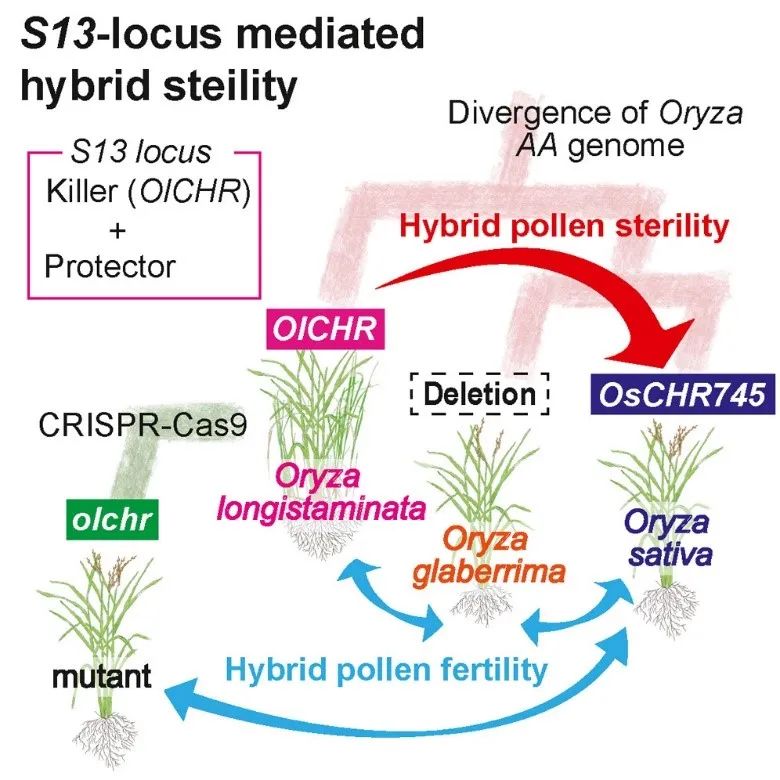
iScience | 染色质重塑因子OICHR是一个调控不同稻种生殖隔离的重要基因 2024年4月23日 0条评论 232次阅读 0人点赞 关于水稻的生殖隔离遗传机制已经得到了广泛的研究,然而,关于不同物种间相关基因的研究则相对较少。在此,研究人员在远缘杂交稻种O. longistaminata和O. sativa中发现了一个表现出自私行为的基因。位于S13位点的自私等位基因S13l损害了雄性育性,在杂合子(S13s/S13l)中特异性地消除了携带O. sativa来源的S13s等位基因的花粉。遗传分析表明,一个编码染色体重塑因子(CHR)的基因参与了这一现象,而O. sativa的多个品种拥有一个截断的基因OsCHR745,而在O. longistaminata中,其同源物OlCHR具有完整的结构。通过CRISPR-Cas9技术介导的功能缺失突变体恢复了杂交稻的育性。值得注意的是,非洲栽培稻自然缺乏OlCHR的同源物,因此能够与携带S13s和S13l的个体兼容。这些结果表明,OlCHR是一个杀手基因,它导致了生殖隔离。综上所述,这项研究揭示了不同稻种间生殖隔离的一个新机制,即通过自私基因OlCHR的作用,使得杂交后代中特定等位基因被排除,从而维持了物种的遗传隔离。这为理解物种形成和演化的遗传基础提供了新的视角。 认识物种的起源是研究生物学的一个基本问题,生殖隔离会干扰两个生物物种之间的基因交流,在各种生殖隔离机制中,合子后隔离屏障已被广泛研究。由于导致杂交不相容的基因在合子或配子产生时降低了其适应性,因此这样的基因是如何进化和在物种内固定下来而不陷入适应性低谷的,一直是一个争论已久的问题。由自私的遗传因素引起的内部遗传冲突被认为是导致杂交不相容的进化力量之一。自私的遗传因素可能由于其“自私的本性”而在群体中迅速固定下来,但也可能被抑制以减少或消除其有害影响。如果这个群体与另一个没有经历过冲突的群体杂交,当自私的遗传因子从抑制因子中分离出来时,就会发生杂交不相容,从而引发杂交冲突。 近期研究揭示了水稻中导致杂交不育基因的自私特性的遗传特征。这些基因通常由两个紧密连锁的组分组成,它们分别作为“杀手”和“保护者”(或“毒素”和“解药”)发挥作用。这种排列是基于最初在Mus musculus的t-单倍型中描述的两组分杂交不育系统。例如,在最近发现的雄性配子不育位点RHS12中,存在两个基因(iORF3/DUYAO和iORF4/JIEYAO)。DUYAO编码一种毒素,它与OsCOX11相互作用,导致线粒体功能障碍和细胞毒性。同时,JIEYAO编码一种解药,通过抑制DUYAO与OsCOX11的相互作用来保护携带它的雄性配子。这种杀手-保护者系统也在亚洲栽培稻中由S5位点介导的杂交不育现象中被发现。此外,在这种物种中,另一种具有自私特性的杂交不育基因的遗传机制涉及杀手-响应者系统(最初在果蝇(Drosophila melanogaster)的SD系统中讨论过;Merrill等人)。例如,杂交不育位点Sa包含两个相邻的基因,SaM(SaM+和SaM-等位基因)和SaF(SaF-等位基因)。在杂交植物中,SaM+和SaF+蛋白共同形成一个杀手,它针对继承SaM-(响应者)位点的花粉。这些发现为我们理解水稻中杂交不育的遗传基础和进化机制提供了新的视角。 S13位点能够诱导杂交雄性不育和雄性特有的转录调控失活(TRD) T65(O. sativa)和W1618(O. longistaminata,如图S1)杂交产生的F1代花粉育性大约为2.3%(如表S4)。为了克隆杂交雄性不育基因,我们将杂合子F1连续回交至轮回亲本T65。随着连续回交至T65,BCnF1的花粉育性逐渐提高(如表S4)。在BC4F1之前的所有回交中,T65一直作为父本,而花粉半不育的BCnF1作为母本。尽管持续回交,但在BC4之后花粉育性仍然保持较低水平,而花粉半不育的分离株种子育性增加,花粉半不育的BC7F1植株产生了完全可育的种子。这些观察结果表明,花粉半不育的遗传因子对种子育性没有产生任何不利影响。从BC7F1自交产生的BC7F2分离株中,我们选出了完全花粉可育的植株。这些植株及其自交后代被视为近等基因系(NIL),命名为T65A+S13l。T65A+S13l幼苗中的花青素色素沉着证实了位于水稻1号染色体上A位点周围的染色体区域被导入(详情见材料与方法)。除了穗数之外,NIL和T65的农艺性状大多相似(如图S2)。 为了确认花粉半不育基因的作用,我们将T65A+S13l与T65进行杂交,并比较了F1代与其亲本的表型。尽管F1代与亲本(T65A+S13l和T65)在整体植株形态和种子育性上没有差异(图1A–F),但F1代的花粉育性显著降低(图1J)。在开花前的花药中,F1代中存在败育的花粉粒,而亲本中几乎所有的花粉粒都正常发育(图1G–I)。在F1植株自交产生的F2群体中,一半植株表现出花粉半不育(花粉育性为45%–55%),另一半植株的花粉育性则≥85%(图1K)。这表明花粉半不育与可育状态之间存在一致且明确的差异;花粉半不育植株和可育植株以1:1的比例分离(图1K)。在通过以T65为父本、F1植株为母本杂交产生的BC1F1植株中,花粉半不育植株和可育植株也以1:1的比例分离,而通过反向杂交产生的BC1F1植株则大多半不育(图1K)。这些结果表明,花粉半不育只发生在杂合子(杂交种)中,且F1杂合子中来自T65的等位基因的雄配子优先败育。因此,我们将这种杂交不育位点命名为S13,并将来自T65和W1618的等位基因分别命名为S13s和S13l。基于上述结果,S13位点的遗传作用被认为是导致mTRD18的花粉杀手类型。在这种情况下,F1杂合子预期在F2群体中以1:1的比例产生S13l纯合子和杂合子(S13s/S13l)。我们使用与A位点连锁的DNA标记E2403B进行验证,并确认在F2群体中,分别有46.6%和50.0%的植株为纯合子(S13l/S13l)和杂合子(S13s/S13l)(图1L)。这些结果表明,来自W1618的S13l等位基因是一个自私的遗传元件(图1M),而S13位点本身则具有调控花粉育性的特殊功能。 在包含102株植物的F2群体中,我们观察到了花粉半不育和可育状态之间的明显区别,且这两种状态以1:1的比例分离(图1K)。这一结果进一步验证了S13位点对花粉育性的控制作用,并表明该位点在遗传过程中遵循了典型的孟德尔遗传规律。花粉半不育状态的植株和可育状态的植株在数量上相等,这支持了我们的假设,即花粉半不育是由杂合子中的一个特定等位基因(S13l)导致的,并且这个等位基因在遗传过程中以显性或半显性的方式表达。为了进一步研究从W1618导入到近等基因系T65A+S13l中的片段长度,我们使用了另外10个DNA标记进行分析。尽管T65和原始野生稻品系W1618之间存在多态性,但在标记S13_i和S13_j处,T65和T65A+S13l具有相同的基因型(图1N)。这一发现表明,在导入的片段中,这些标记所代表的区域在T65和T65A+S13l之间是保守的,没有发生变异。这些结果对于理解S13位点的遗传结构和功能具有重要意义。首先,它们证实了S13位点确实位于从W1618导入到T65A+S13l的片段内。其次,通过比较不同标记的基因型,我们可以进一步缩小S13位点所在的具体区域,为后续的图位克隆和功能分析提供重要线索。 CRISPR-Cas9介导的突变确实恢复了杂交雄性生育能力 首先假设在T65中,OsCHR745基因的过早终止密码子会产生一个截短且功能失调的蛋白质,而OlCHR则产生功能正常的蛋白质;这种变化会导致S13s和S13l等位基因具有不同的功能(图2A)。为了验证这一假设,我们在T65A+S13l中开发了针对OlCHR的CRISPR-Cas9突变体。我们设计的引导RNA能够识别T65A+S13l中OlCHR第一个外显子上的特定目标。然后,我们产生了在PAM序列上游分别有2个和1个碱基缺失的突变体,分别命名为olchr_1和olchr_2(图2B–D)。我们将这些突变体与A58(一种O. sativa品种)进行杂交。A58在A位点上没有显示杂交花粉不育和紧密连锁的S13位点的TRD,当它与T65杂交时,在F2群体中也是如此;而当它与T65S13l杂交时,则显示出杂交花粉不育和TRD(图S8)。这一结果表明,A58在S13位点上具有S13s等位基因。我们还确认,T65和A58之间候选基因的编码区没有单核苷酸多态性(SNPs)和插入/缺失(InDels),因此我们使用这种品种作为S13s携带者的代表。A58与olchr_1和A58与olchr_2之间F1杂交后代的花粉育性分别为87.6%和86.6%(图2E和F)。这些值与T65和T65A+S13l的花粉育性没有显著差异(图2F),表明在这些杂交组合中没有发生杂交雄性不育。因此,OlCHR可能与S13位点介导的杂交不育有关。在S13位点介导的杂交不育系统中,存在通过雄性配子流产来优先传递S13l等位基因的情况(S13s/S13l)。因此,我们检查了由F1植物衍生的F2群体的分离模式,以确认是否发生了S13l等位基因的优先传递。在F2群体中,分离模式与孟德尔1:2:1比例没有显著差异(图2G),表明在F2群体中没有发生mTRD。这些发现进一步支持了我们的假设,即OlCHR在S13位点介导的杂交不育中扮演了重要角色。通过CRISPR-Cas9技术编辑OlCHR基因,我们成功地恢复了杂交雄性生育能力,这为我们理解该基因在杂交不育中的具体作用提供了直接证据。此外,对F2群体分离模式的分析也表明,S13l等位基因并没有通过优先传递的方式影响群体的遗传结构。这些结果不仅有助于我们深入理解水稻中的遗传不育机制,也为未来通过基因编辑手段改良作物品种提供了有价值的参考。 广泛存在的终止密码子等位基因 在上述分析中,我们发现OlCHR及其同系物OsCHR745中的过早终止密码子导致S13位点等位基因的功能改变。其中,五个单倍型(Hap.6、Hap.23、Hap.24、Hap.28和Hap.30)包含超过10个品种(图3A和附图S9)。对于另外20个单倍型,每个单倍型仅包含一个品种(附图S9)。在所有单倍型中,有八个具有提前终止密码子(TAA),即在1号染色体25,527,848 bp位置具有胸腺嘧啶。具有这种终止密码子的品种频率几乎占到了O. sativa种群的四分之一。我们在Hap. 28和Hap. 14中分别发现了另外两个SNP,它们分别在OsCHR745的第5个和第20个外显子上导致提前终止密码子,这表明在这些单倍型中OsCHR745也是非功能的(图3A)。为了考察单倍型与O. sativa亚群之间的关系,我们根据每个品种所属的亚群对每个单倍型中的品种进行了分类(图3B和C,每个亚群的名称基于Rice SNP-Seek数据库)。这一分析揭示了亚群特有的单倍型分布。对于Hap. 23和24,亚热带和温带品种分别广泛分布(图3B)。这些发现表明,OsCHR745基因的提前终止密码子变异在O. sativa的不同亚群和品种中广泛存在,且具有一定的分布规律。这种分布模式可能与不同亚群的生态适应性、遗传背景以及进化历程有关。 在本研究中,我们利用CRISPR-Cas9进行突变体筛选,在杂种不育位点开发了一个非功能性的“Killer”等位基因,该等位基因不会引发由S13位点介导的杂种不育。并鉴定了到了一个在两个水稻物种杂交不育系统中的“Killer”功能基因—OlCHR基因。为了全面描述这一系统,未来的研究应旨在鉴定“Protector”或“Responder”基因。由于OlCHR与编码染色质重塑因子的基因具有同源性,未来的研究应探索染色质状态与杂种雄性不育之间的关系。 文章来源:植物生物技术Pbj < 上一篇 下一篇 > 发表回复 取消回复您的电子邮箱地址不会被公开。 必填项已用*标注 * *