在植物的绚烂生命篇章中,隐秘且错综复杂的微观防御战斗不断地展开。这其中,固着生物与它们的微生物对手进行着持续的军备竞赛。作为陆地生态系统的基石,植物已经进化出一系列复杂的防御机制来抵御病原体的无情攻击。植物和环境之间的这种动态相互作用证明了双方的进化能力,植物发展出包括PTI和ETI在内的多层免疫系统(图1)。这篇综述旨在提供对植物内源性免疫逐步激活的当前知识的全面概述,重点关注PRRs和NLRs分别识别细胞外和细胞内效应子的过程,强调了这些途径之间复杂的相互作用及其对整体植物免疫反应的贡献。
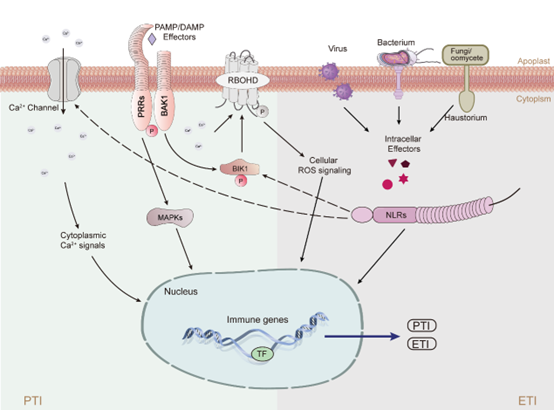 图1. 植物免疫信号网络模式图
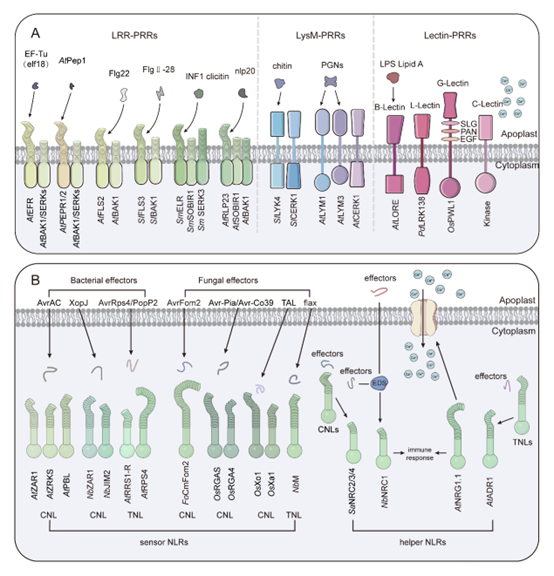
植物的第一道免疫防线由细胞表面的模式识别受体(PRRs)构成,它们通过招募共受体来识别入侵病原体的保守特征,包括病原体相关分子模式(PAMPs)或宿主衍生的损伤相关分子模式(DAMPs),从而激活模式触发免疫(PTI),为植物提供基础的疾病抵抗力。PRRs包括受体样激酶(RLKs)和受体样蛋白(RLPs),它们具有识别多种配体的功能域或基序,如类固醇、多肽、多糖、脂多糖以及病原细菌释放的效应子,还有宿主的内源性分子,这些分子能有效增强植物疾病抵抗力(图2)。
与此同时,植物也演化出了核苷酸结合/亮氨酸富集重复(NLR)受体来对抗这些逃避策略。NLRs直接结合效应子或在宿主细胞内感知其存在,触发被称为效应子触发免疫(ETI)的第二层防御。植物NLRs由抗性(R)基因编码,具有包含保守中心核苷酸结合域(NB-ARC)和C末端LRR域的多域结构。它们根据N末端域的不同被分类为TIR结构域或Frizzled结构域,进一步细分为G10型CC-NLRs、RPW8型CC-NLRs和Rx型CC-NLRs。LRR域负责直接或间接识别效应子,NB-ARC域具有调节NLR激活的ATG结合活性,而N末端域参与NLR激活后的下游信号转导(图2)。
图2. PRRs和NLRs介导的免疫信号传导
植物免疫的PTI-ETI交互作用展示了一个综合性的防御体系,其中PRR和NLR这两类受体虽然分别触发,却被认为在植物的统一防御系统中扮演着相互依赖的角色(图3)。这种交互作用的亮点在于,NLR介导的免疫反应通常与PTI共同激活,或者在PTI的基础上进一步发展成ETI,而独立于PTI之外激活NLR的情况较为罕见。这种机制的创新之处在于,PRR和NLR之间的相互作用能够促进细胞间信号的传递,从而激活一系列下游防御基因。这种复杂的信号网络不仅增强了植物对病原体的抵抗力,而且允许植物在面临环境变化时快速调整其防御策略,同时有效利用资源。通过这种方式,植物能够在保持生长和发育的同时,有效地抵御病原体的侵袭。
图3. PRRs和NLRs介导免疫系统的串扰模型
在植物免疫的PTI和ETI的协同信号转导中,活性氧(ROS)和钙离子(Ca2+)信号发挥着至关重要的作用(图4,5)。这两种免疫机制在不同的时间和空间范围内运作,需要借助第二信使作为“语言代码”,将外部信号传递到细胞内部,并激活下游的信号网络。越来越多的证据强调了ROS和Ca2+信号在精细调节植物免疫反应中的细胞网络中的重要性。
图4. 植物免疫中ROS的生成和清除通路示意图
图1. 植物免疫信号网络模式图
植物的第一道免疫防线由细胞表面的模式识别受体(PRRs)构成,它们通过招募共受体来识别入侵病原体的保守特征,包括病原体相关分子模式(PAMPs)或宿主衍生的损伤相关分子模式(DAMPs),从而激活模式触发免疫(PTI),为植物提供基础的疾病抵抗力。PRRs包括受体样激酶(RLKs)和受体样蛋白(RLPs),它们具有识别多种配体的功能域或基序,如类固醇、多肽、多糖、脂多糖以及病原细菌释放的效应子,还有宿主的内源性分子,这些分子能有效增强植物疾病抵抗力(图2)。
与此同时,植物也演化出了核苷酸结合/亮氨酸富集重复(NLR)受体来对抗这些逃避策略。NLRs直接结合效应子或在宿主细胞内感知其存在,触发被称为效应子触发免疫(ETI)的第二层防御。植物NLRs由抗性(R)基因编码,具有包含保守中心核苷酸结合域(NB-ARC)和C末端LRR域的多域结构。它们根据N末端域的不同被分类为TIR结构域或Frizzled结构域,进一步细分为G10型CC-NLRs、RPW8型CC-NLRs和Rx型CC-NLRs。LRR域负责直接或间接识别效应子,NB-ARC域具有调节NLR激活的ATG结合活性,而N末端域参与NLR激活后的下游信号转导(图2)。
图2. PRRs和NLRs介导的免疫信号传导
植物免疫的PTI-ETI交互作用展示了一个综合性的防御体系,其中PRR和NLR这两类受体虽然分别触发,却被认为在植物的统一防御系统中扮演着相互依赖的角色(图3)。这种交互作用的亮点在于,NLR介导的免疫反应通常与PTI共同激活,或者在PTI的基础上进一步发展成ETI,而独立于PTI之外激活NLR的情况较为罕见。这种机制的创新之处在于,PRR和NLR之间的相互作用能够促进细胞间信号的传递,从而激活一系列下游防御基因。这种复杂的信号网络不仅增强了植物对病原体的抵抗力,而且允许植物在面临环境变化时快速调整其防御策略,同时有效利用资源。通过这种方式,植物能够在保持生长和发育的同时,有效地抵御病原体的侵袭。
图3. PRRs和NLRs介导免疫系统的串扰模型
在植物免疫的PTI和ETI的协同信号转导中,活性氧(ROS)和钙离子(Ca2+)信号发挥着至关重要的作用(图4,5)。这两种免疫机制在不同的时间和空间范围内运作,需要借助第二信使作为“语言代码”,将外部信号传递到细胞内部,并激活下游的信号网络。越来越多的证据强调了ROS和Ca2+信号在精细调节植物免疫反应中的细胞网络中的重要性。
图4. 植物免疫中ROS的生成和清除通路示意图
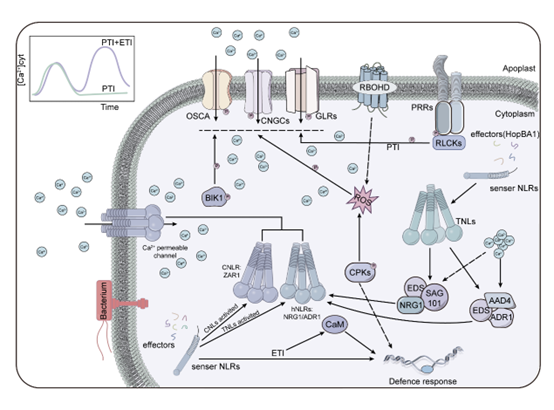
图5. 植物免疫中Ca2+信号传导通路示意图
北京林业大学生物科学与技术学院博士研究生于晓倩为论文第一作者,夏新莉教授为本文通讯作者。此外,团队硕士研究生牛浩强以及青年教师刘超、王厚领参与了该研究工作。该论文得到农业生物育种重大项目(2022ZD0401503)和国家自然科学基金(32071734)的资助。