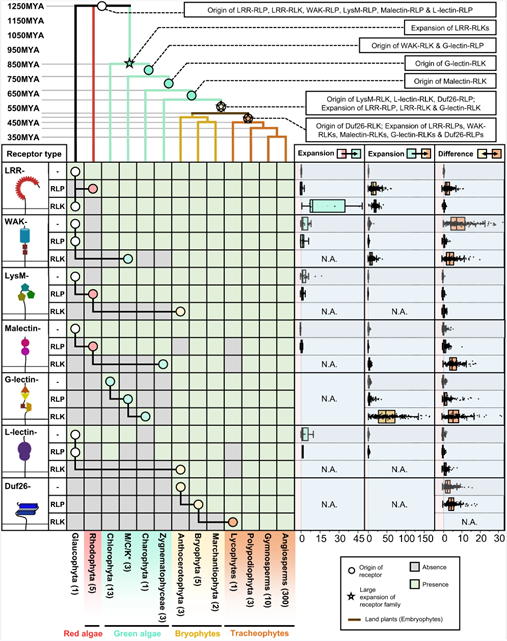
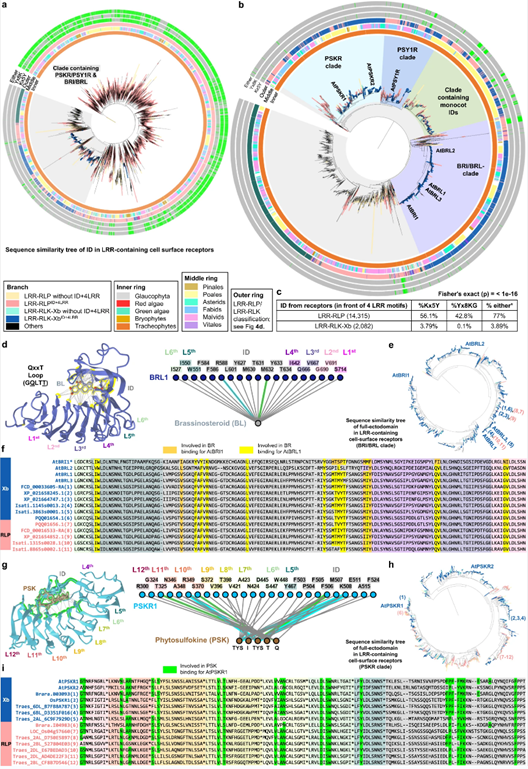
图4. 植物中LRRID+4LRR的起源和进化。
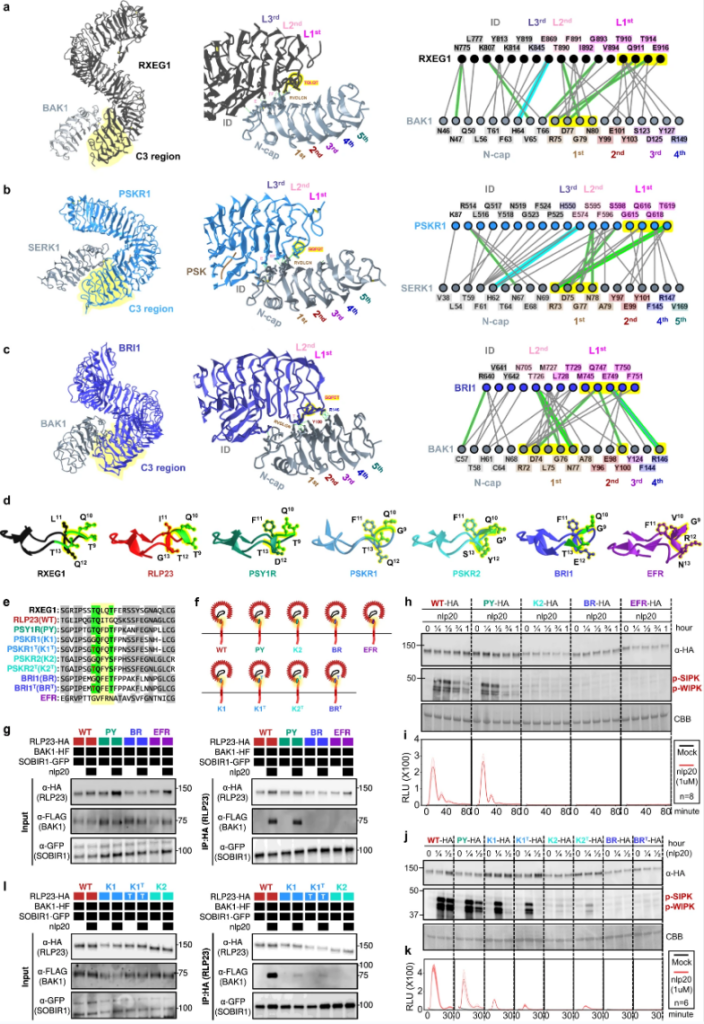
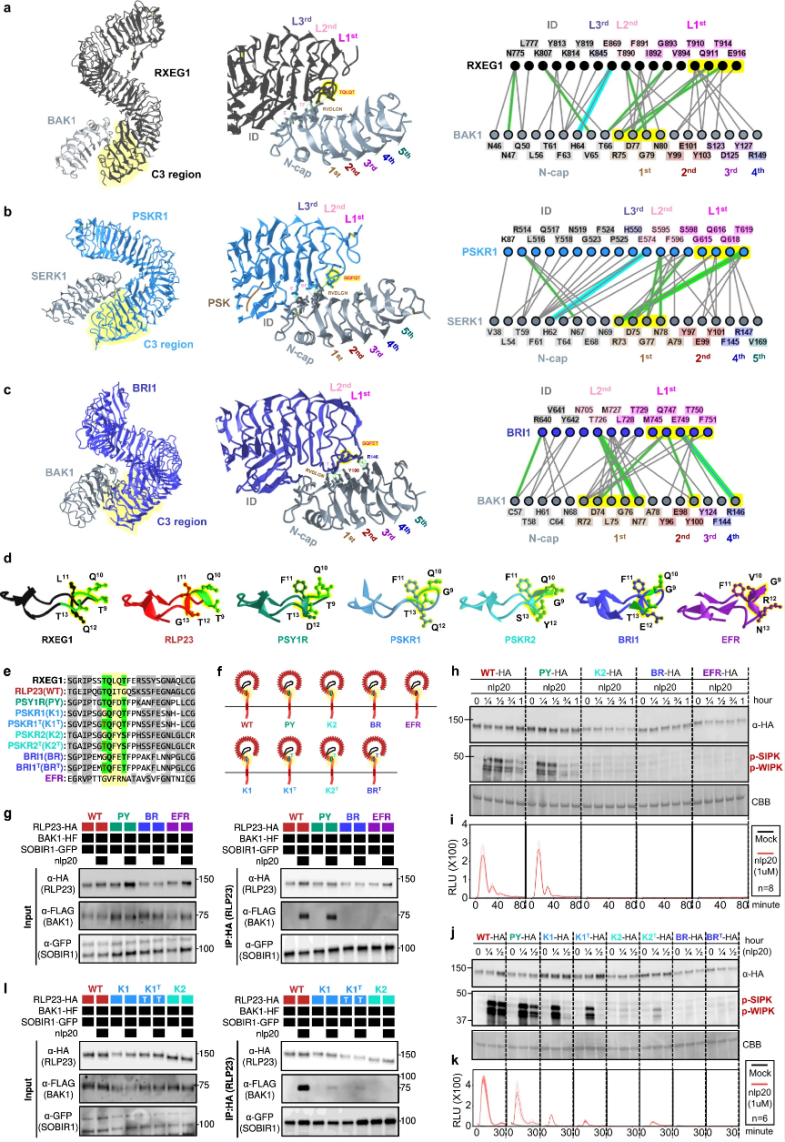
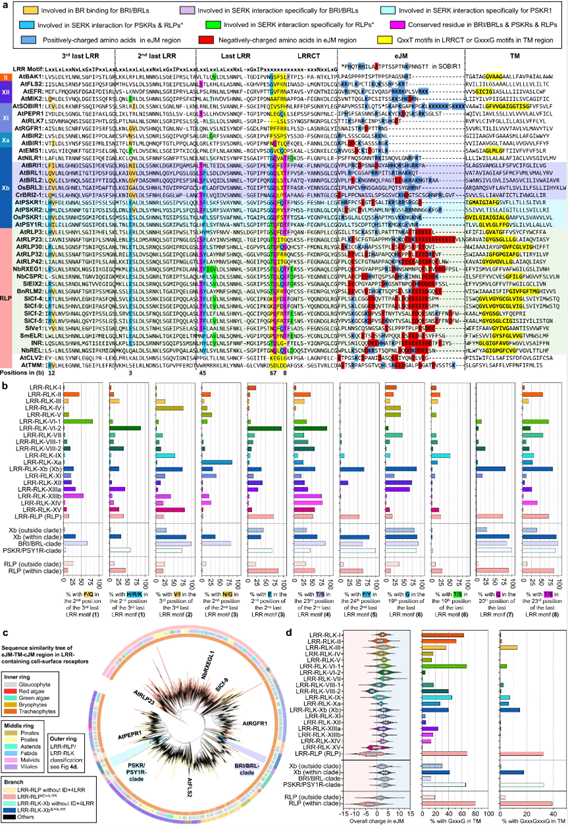
图5. LRR-RLP和LRR-RLK-Xb中C3区域的功能特征。
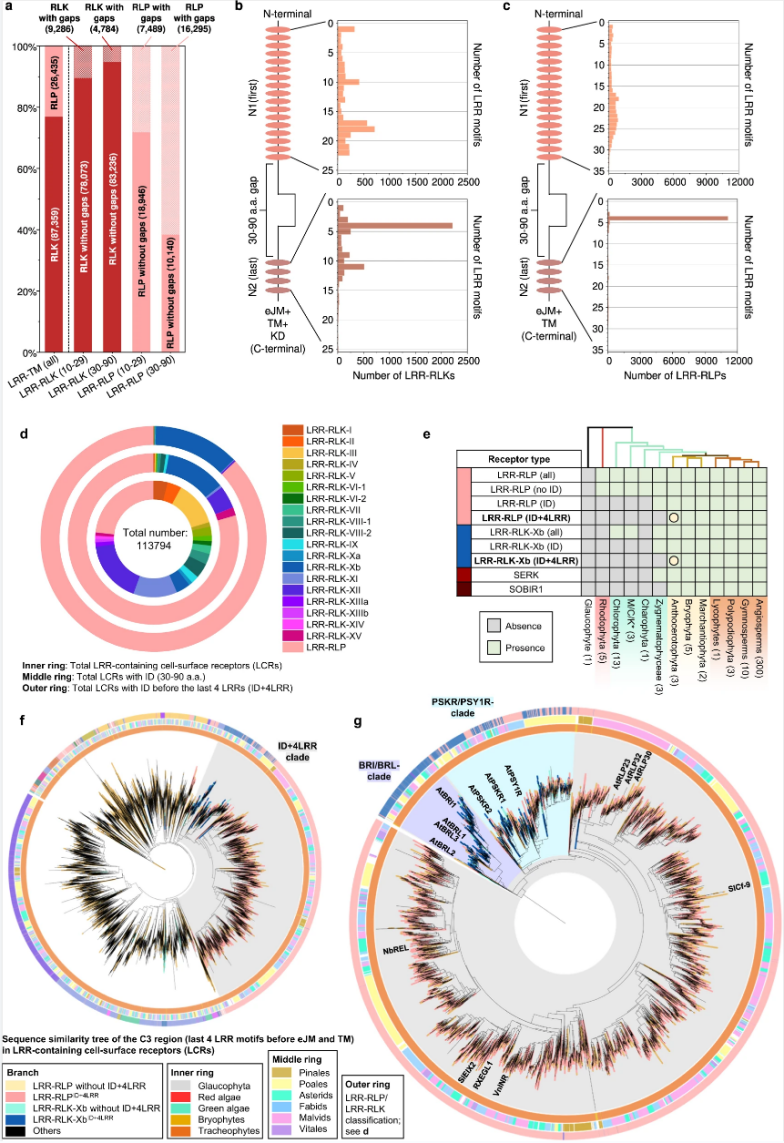
为了剖析LRR-RLP和LRR-RLK-Xb的胞外域起源,他们研究了这两组受体之间IDs的相关性。结果表明LRR-RLK-Xb和LRR-RLP的IDs可能起源于共同的祖先,在LRR-RLK-Xb和LRR-RLP分歧后,LRR-RLP的IDs不断扩展和多样化。
图6. LRR-RLPs和LRR-RLK-Xbs的起源。
LRR-RLP和LRR-RLK-Xb胞外域的共同起源,这些受体在出现分歧后必定在免疫和发育过程中经历了专门化。研究人员发现LRR-RLP和PSYR/PSY1R进化枝的TM区域也可能具有共同的起源。
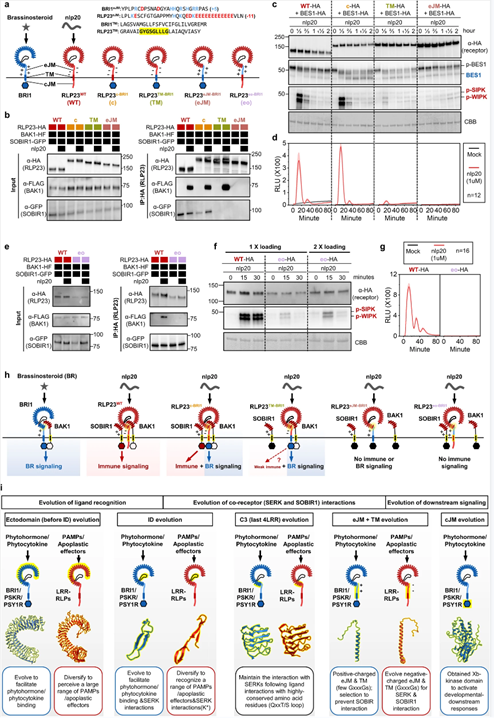
他们生成了eJM、TM以及BRI1和RLP23的胞质区域的多个嵌合体,评估LRR-RLK-Xb和LRR-RLP中eJM-TM胞质区域的功能。实验结果表明,LRR-RLP的eJM区域对于蛋白质积累以及配体感知时与BAK1的相互作用非常重要。LRR-RLP和LRR-RLK-Xb的eJM、TM和胞质区域确实专门用于将不同的蛋白质招募到受体复合物中,这使得它们能够激活不同的下游反应。
图7.LRR-RLK和LRR-RLP中末端4个C3、eJM和TM区域的排列和特征。
图8. LRRID+4LRR对差异下游信号通路的适应。
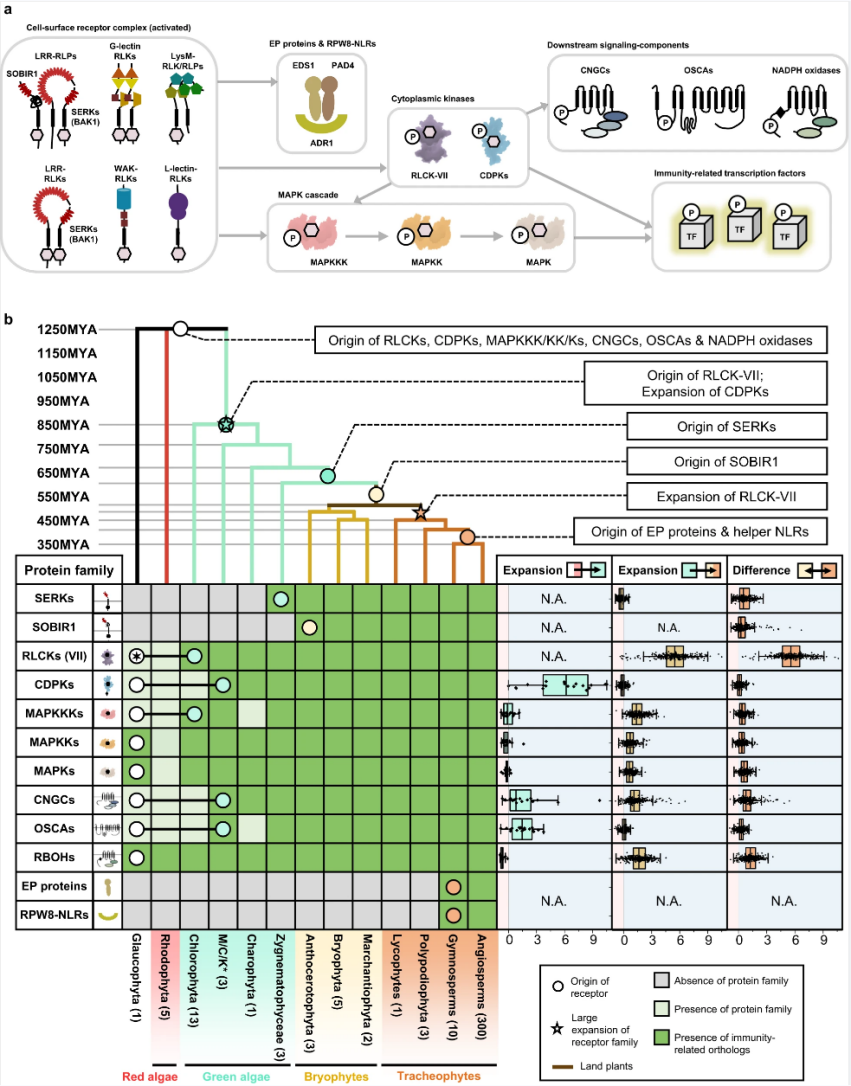
研究人员工作揭示了细胞表面受体促进植物适应的多种进化机制:1. 体数量及其识别特异性的扩展使植物能够识别更广泛的特定于某些胁迫和环境信号的分子。2. 信号网络的发展日益复杂似乎增强了差异响应的灵活性和调节能力,以跟上快速变化的环境。3. 现有受体对特定信号的适应,LRR-RLP和LRR-RLK-Xb之间的结构相似性意味着免疫特异性细胞表面受体和发育特异性细胞表面受体之间的共同起源。