
先前的研究表明,质体内容物可以从细胞器中逸出,这表明质体转基因技术可能在细胞的其它位置发挥作用。为了验证这一假设,研究者们创造了烟草质体转化体,其表达核编码的八氢番茄红素去饱和酶(PDS)基因的片段,如果RNA逃逸到细胞质中,该基因能够催化转录后基因的沉默。研究者们在叶绿体基因组中插入了一个dsRNA,该转基因携带编码PDS的核基因片段,而PDS是类胡萝卜素和叶绿素的积累所必需的。叶绿体中携带这种转基因的植物表现出白色的“漂白”表型,这就表明在植物发育的不同阶段,类胡萝卜素和叶绿素积累的很少或者说是几乎没有积累。这也能够表明叶绿体表达的dsRNA转基因可以沉默核基因的表达。出乎意料的是,研究者们发现质体表达的dsRNA在细胞质中产生了21个核苷酸的长度较短的小干扰RNA。更为有趣的是,质体表达的dsRNA通过小RNA生物途径引起基因沉默,这其中包括“未触发的siRNA”。研究者们下一步则想了解从叶绿体基因组表达的dsRNA是如何进入细胞质并绕过了PhasiRNA途径的初始生物合成步骤。
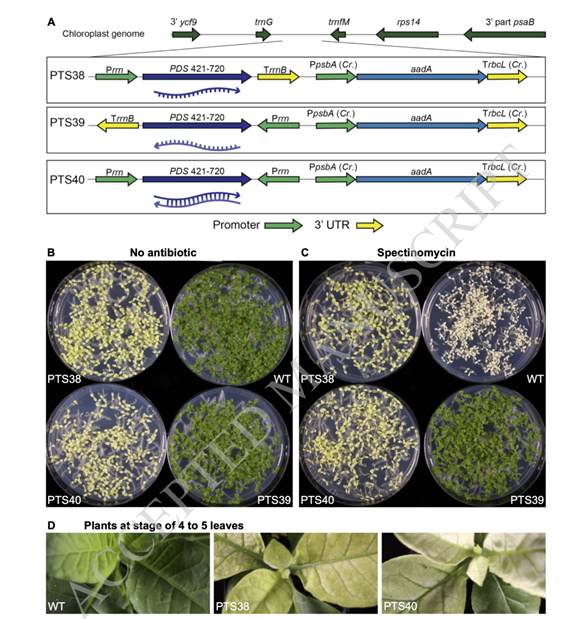
研究者们为了证实质体编码性状的母体遗传,分析了自受精转成细胞系幼苗的抗性及其色素缺乏表型。正如预期的那样,野生型幼苗在缺乏抗生素的培养基上是绿色的(图1B),但在含有壮观霉素的培养基(图1C)上被均匀地漂白成白色,表明它们对抗生素的敏感性。相反,来自PTS38和PTS40系自受精的幼苗具有中间表型,在含有或不含有壮观霉素的平板上生长时,均匀地漂白为黄色而不是白色(图1B-C)。这些结果表明,PTS38和PTS40品系对抗生素和细胞质具有抗性,而所导致的色素缺乏可能会引起PDS基因的沉默。
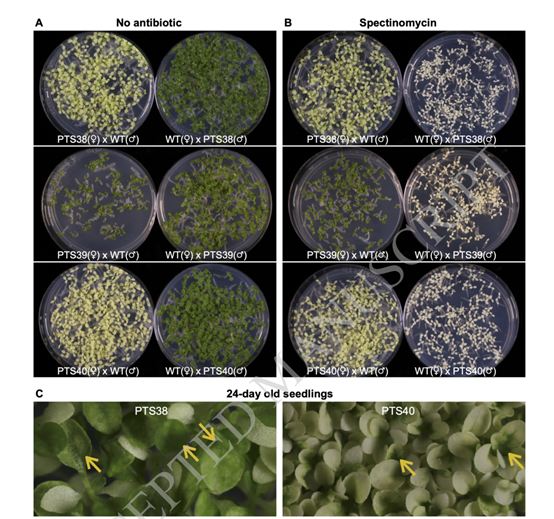
研究者们利用质体转化系与野生型植物的相互杂交来测试质体转基因性状的母体遗传。当PTS38和PTS40植物作为雌性亲本与野生型植物杂交时,在缺乏抗生素的培养基上,所有幼苗都一致缺乏色素(图2A)。相反,当质体转化系在与野生型植物的杂交中用作雄性花粉供体时,所有幼苗都是均匀的绿色(图2A)。这些结果证实在PTS40和PTS38系的T2和T3代,核基因组中不存在PDS转基因的活性拷贝。
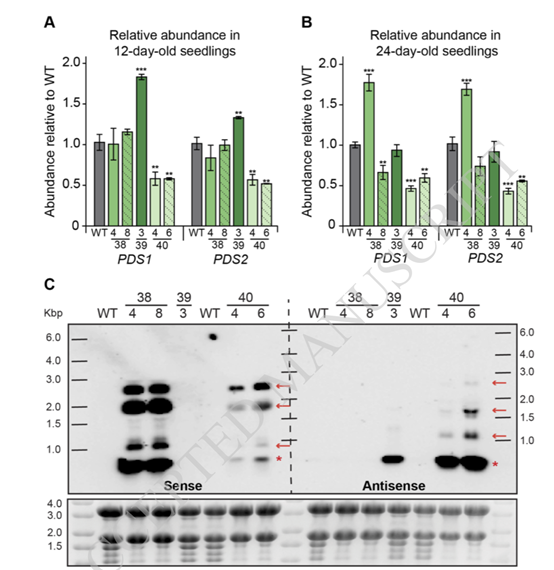
研究者们为了确定质体转化系的色素缺乏是否是由于细胞核编码的细胞质定位的PDS1和/或PDS2信使核糖核酸的沉默,使用RT-qPCR检测了T1转基因和野生型幼苗中PDS1 和PDS2的丰度。这证实了在PTS40品系中观察到的漂白表型可能是由于核编码的PDS mRNA水平的敲低所导致的,而在PTS38品系中观测到的表型则可能是因为其它不同的机制所导致的。
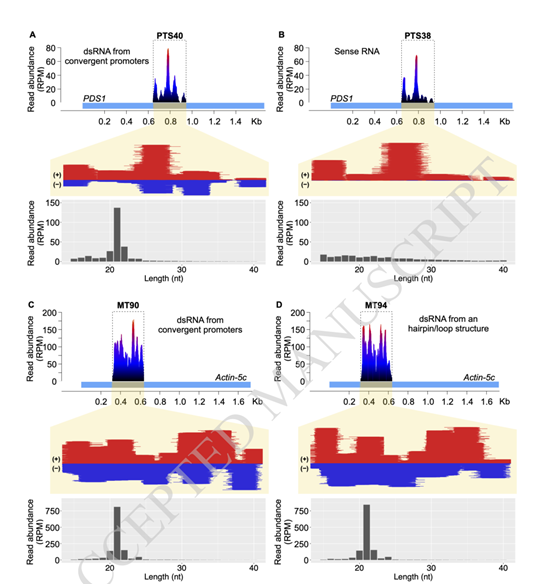
综上所述,在这项研究中,研究者们测试了质体表达的转基因RNA是否可以逃离细胞器,并在细胞质转录后基因沉默途径中发挥作用。结果表明,常见的RNA从质体逃逸到细胞质的过程,可用于敲除宿主核编码的基因,而该项研究也通过质体转化技术扩展了生物技术工具。
文章来源:植物生物技术Pbj
