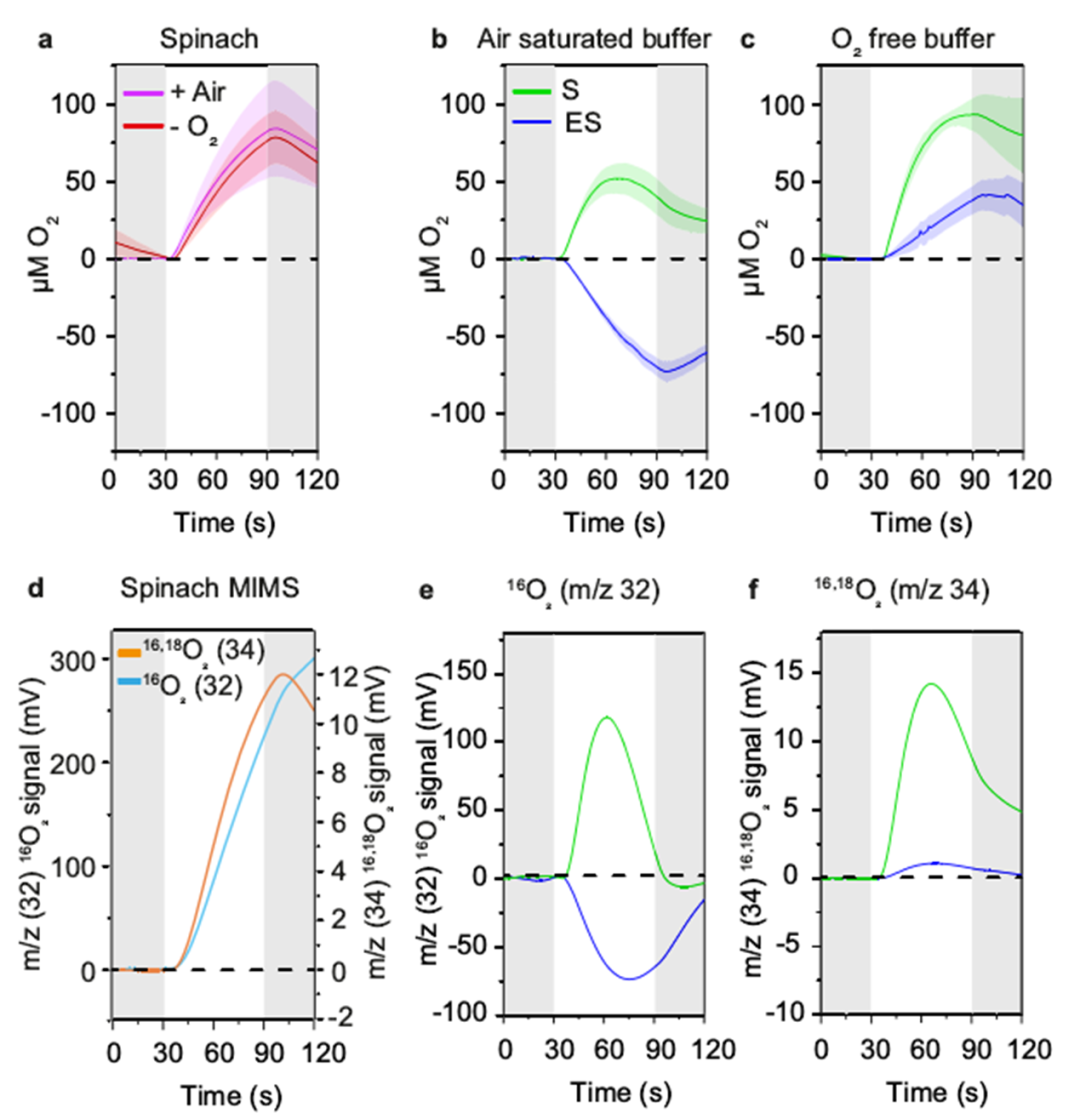
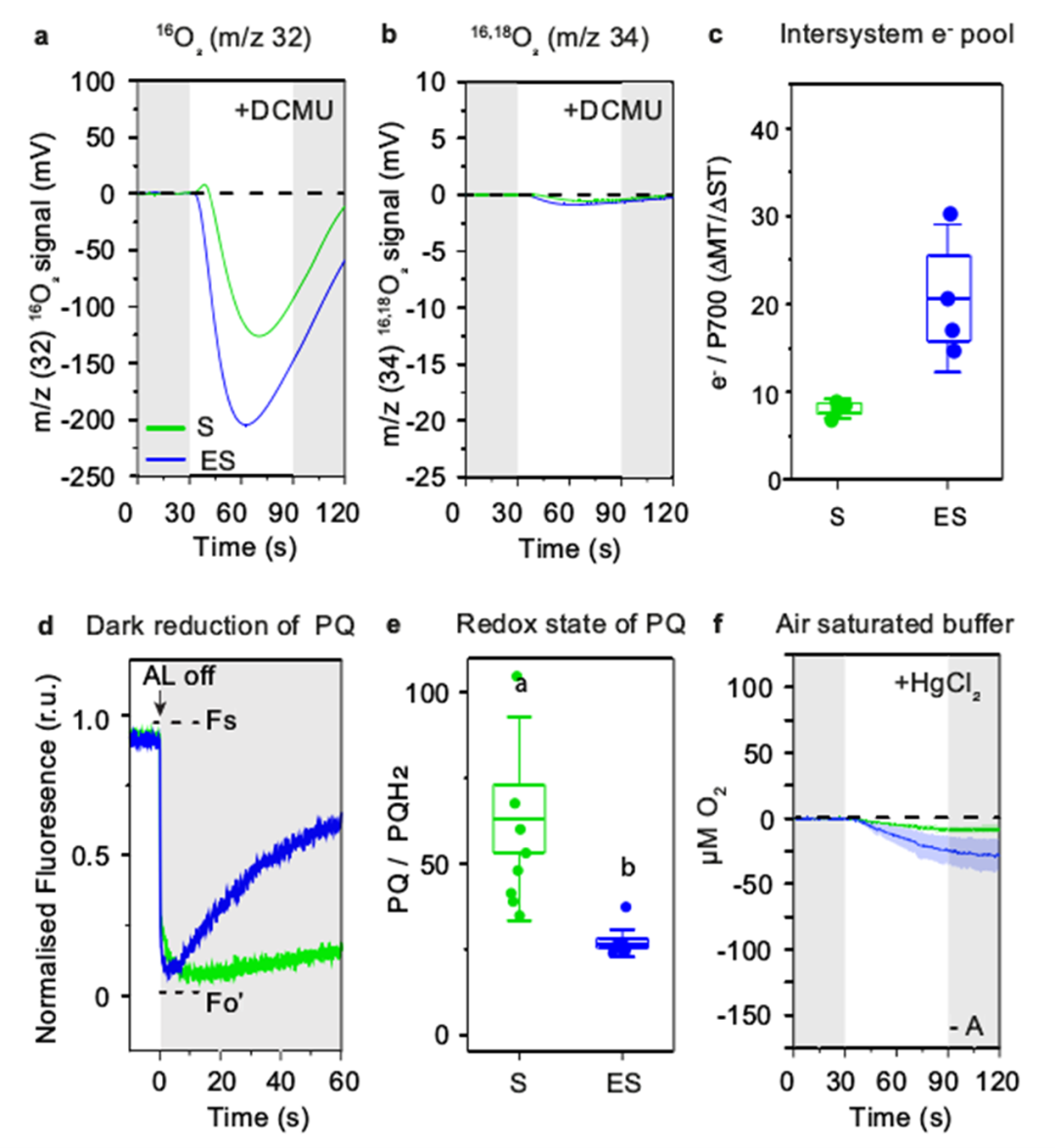
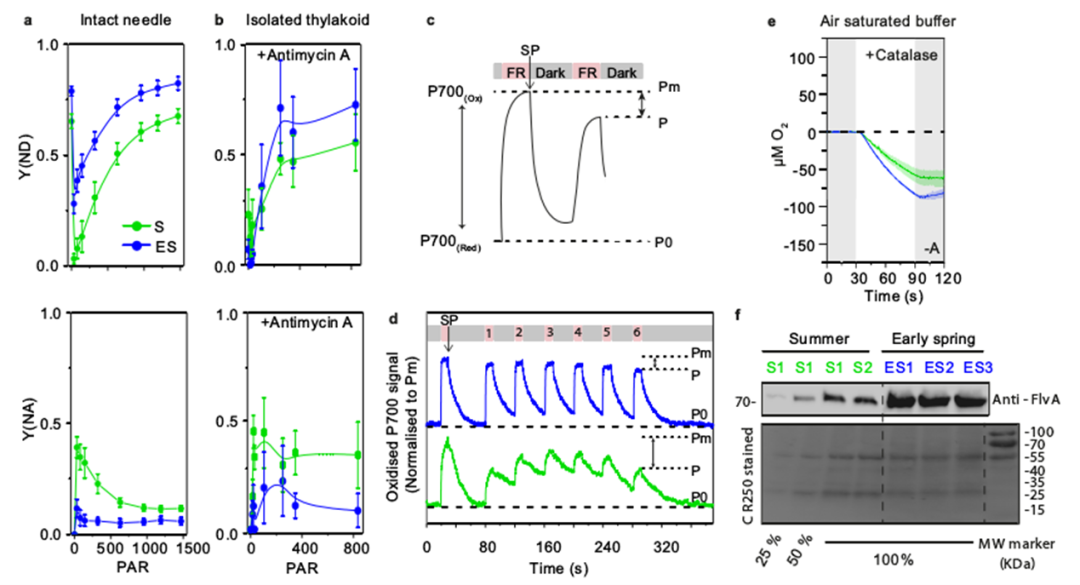
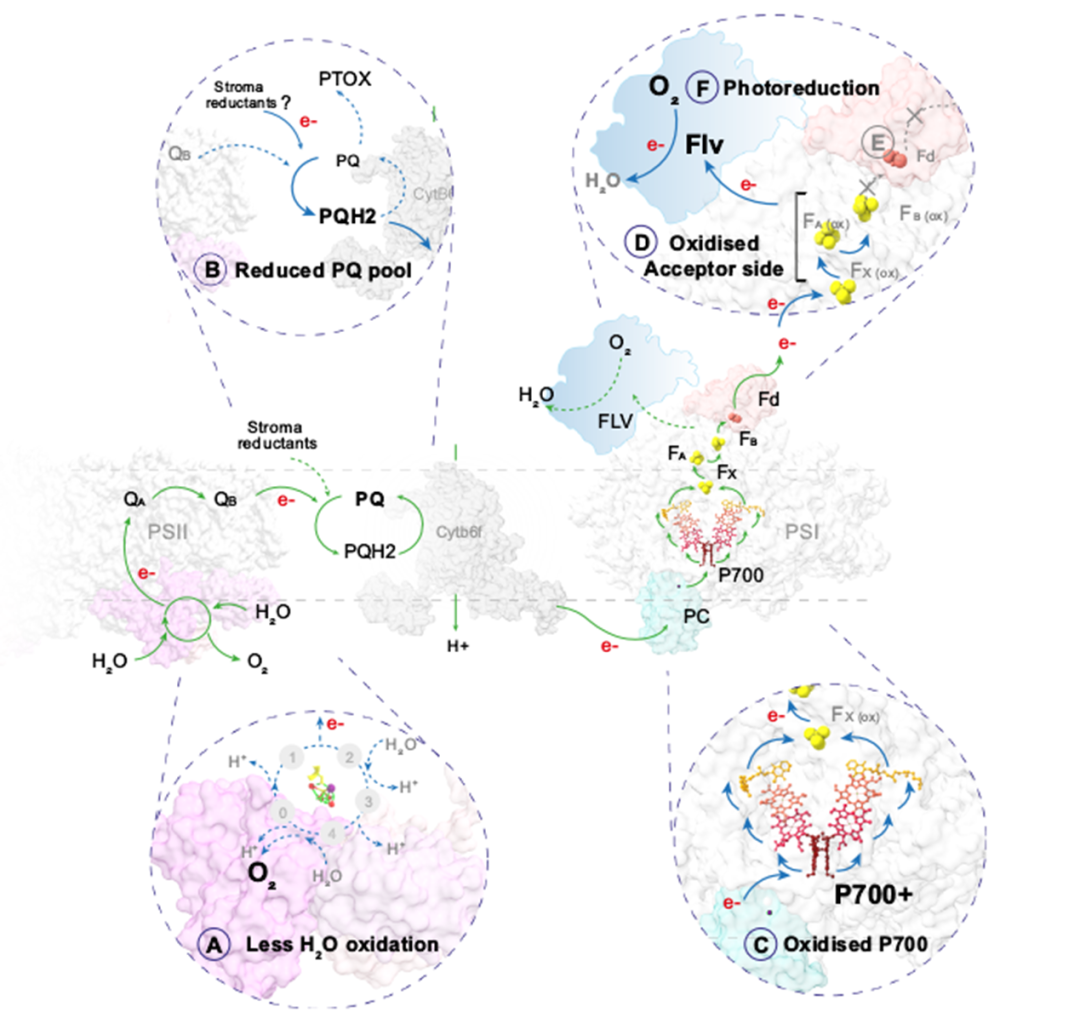
地球大气中的氧气是由生长在水中和陆地上的光合生物产生的。北方森林覆盖了地球森林总面积的33%,为全球碳平衡和氧气生产做出了重大贡献。树木从H2O中提取的电子通过几种氧化还原载体进一步转移到光系统PSⅠ,随后被NADP+接受,在光合电子转移链中产生NADPH3。随后在暗反应中,在卡尔文循环中涉及NADPH的二氧化碳同化。光反应和暗反应之间的氧化还原不平衡通常会导致活性氧(ROS)的形成,从而损害光系统。因此,植物进化出了多种保护机制,如PSII7中的非光化学猝灭(NPQ)、PSⅡ周围的Mehler反应和叶绿体基质中RuBisCO的光呼吸。在光呼吸中,RuBisCO固定O2而不是CO2,并通过间歇的线粒体和过氧化物酶体间穿梭体释放CO2。目前针叶树PSⅠ的保护机制尚不明确。裸子植物中缺乏类囊体NDH复合物(NADH脱氢酶样)。而且在低温下,其光呼吸已被证明不是主要的电子吸收方式。显然,从被子植物的研究中得出的结论可能不适用于针叶树。蓝藻、藻类和苔藓有一个额外的黄二铁(Flv)蛋白介导的途径,利用多余的电子在PSI受体侧将O2直接还原为H2O。最近,类似的机制也被预测存在于裸子植物(包括针叶树)中,但尚未得到实验证实。瑞典尤梅铎大学植物生理学系尤梅铎植物科学中心的Pushan Bag团队联合瑞典尤梅夫大学瑞士生物化学中心化学系及加拿大西安大略大学生物学系的研究人员近日在Nature Communications上发表了一篇题为“Flavodiiron-mediated O2 photoreduction at photosystem I acceptor-side provides photoprotection to conifer thylakoids in early spring”的研究论文。研究表明,即使在早春(ES)极低温度与高太阳辐射同时存在的光照下,苏格兰松(Pinus sylvestris L)和挪威云杉(Picea abies)针叶的绿色类囊体膜也表现出强烈的氧气消耗。通过使用不同的电子传递链抑制剂,发现这种不寻常的光诱导氧消耗发生在光系统(PS)Ⅰ周围,并与ES类囊体中更高丰度的黄二铁(Flv)A蛋白相关。通过P700吸收变化,证明了通过O2光还原从PSⅠ受体侧清除电子是ES的主要替代途径。结合P700吸光度和免疫检测,证明了flv依赖的O2消耗是缓解早春植物在低温和高辐射联合胁迫下基质代谢还原剂对PQ池过度还原的主要功能电子汇,为维管植物在PSI周围的这种光还原O2及其季节变化提供了功能证据。维管植物的这种光保护机制表明,针叶树已经形成了适应恶劣环境生长的进化轨迹。克拉克电极O2测量表明,在菠菜类囊体中,O2产量与培养基中的O2水平无关,因为在空气饱和和无O2缓冲液中O2交换相似。相比之下,S松树类囊体膜中的O2进化强烈依赖于培养基中的O2水平。在空气饱和缓冲液中,松类囊体的氧演化约为菠菜类囊体的一半。此外,O2产率在照明周期的前30秒(60秒)达到最大值,然后在接下来的30秒(60~90秒)缓慢下降。在无O2缓冲液中,S类囊体的O2进化在前50秒增加,后10秒趋于平稳。这一数据表明,在体外,S松树类囊体的O2产生受到光失活机制的抑制,该机制依赖于样品试管中的O2水平。有趣的是,在不添加PPBQ和FeCy的空气饱和缓冲液中,菠菜膜没有显示任何氧气交换,而ES和S松类囊体仅显示氧气消耗。PETC中的所有还原反应都需要电子,而电子通常是由PSⅡ中的水氧化提供的。为了确定参与O2光消耗的电子是否来自PSⅡ,研究人员将DCMU添加到类囊体悬浮液中。在DCMU存在下,16,18O2产率完全降低,这表明PSⅡ中的H2O氧化在S和ES中都被完全阻断。令人惊讶的是,S和ES样品都消耗了16O2,但ES中16O2的负振幅比S强。这表明PSⅡ中的H2O氧化没有为S和ES中的O2光还原提供电子。因此,研究人员得出结论,在松类囊体膜中,同时发生的O2演化和消耗是两个独立的过程,在S和ES中都发生,但大小不同。这种差异导致了S和ES之间的净O2产量模式的差异,并且O2消耗不依赖于PSⅡ。研究发现,与未添加HgCl2相比,S和ES中的O2消耗都减少了。综合来看,这些结果表明,ES样品中的O2消耗是在PSI周围发生的光还原过程,电子来自非光化学还原的PQ池,PTOX不是O2光还原的主要位点。已知PSI周围的氧气消耗对PSⅠ的氧化还原状态有深远的影响。与前人研究一致,在笔者的研究中,与S样品相比,随着辐照度的增加,完整ES针中P700的氧化程度增加(Y[ND]增加),受体侧限制(Y[NA])减少。使用了比以前报道的高3倍的抗霉素A浓度,在这个浓度下,抗霉素A有效地调节P700再还原动力学。然而,研究人员发现SP测量的总体趋势相似,在含有抗霉素a的类囊体和不含抗霉素a的完整针上,即随着辐照度的增加,Y(ND)高于S,Y(NA)低于S。表明ES类囊体的氧气消耗伴随着更高的P700氧化和更高的黄二铁蛋白丰度。在S类囊体膜照射下,H2O分裂成O2产生的电子使PQ降低,并进一步通过Cyt b6f到PC,再到P700。接收到电子后,P700被还原,然后在光照下通过向电子受体(FX到FA到FB)提供电子而被再氧化,这些电子受体随后被Fd吸收进行正向反应。在ES(A)中,尽管H2O分裂反应减慢,但PQ池仍处于高度还原状态(B)(很可能是ndh-2介导的基质还原),这有争议地将电子提供给P700。照明(C), P700容易被氧化的捐赠电子受体(外汇、FA和FB)(D)。因此,该受体(外汇、FA和FB)成为高度降低电子正反应的需求是有限的,由于下调的CBB (E)。然而,外汇,FA和FB仍然处于氧化状态的电子被Flv蛋白质(F)。Flv活动结果强劲的O2消费更高,导致净O2消耗量和PSI donor-side有限。研究人员认为也许flv对针叶树来说是一种有用的机制,当条件变得更糟时,它可以迅速被调用,当条件变得更好时,它可以被拆除。针叶树失去了I型ndh介导的CEF,只保留了Pgr5/Pgrl158。这些不同于被子植物的进化轨迹可能是适应性的,因为针叶树通常适应恶劣的环境,但在富裕的生态系统中,它们与被子植物的竞争通常不那么好。也许Flv蛋白提供了更好的保护,但ndh介导的CEF在更有利的环境下提供了更好的能源经济性。因此,Flvs可能是针叶树“安全总比后悔好”进化策略的一部分。文章来源:植物生物技术Pbj