绿色植物可以通过由核酮糖-1,5-二磷酸羧化酶/氧化酶(Ribulose-1,5-bisphosphate carboxylase/oxygenase,简称Rubisco)介导的光合作用将大气中的二氧化碳转化为碳水化合物。据估计,Rubisco是地球上最丰富的蛋白之一,每年固定大约10.511亿吨的二氧化碳。然而,它的活性受到抑制性糖类(如木糖酮-1,5-二磷酸酯,简称XuBP)的结合而受损。在植物中,XuBP的合成速率远高于其他糖类抑制剂,并且随温度升高而增加。XuBP磷酸酶(CbbY)可将XuBP转化为无害的CBB循环中间产物木糖-5-磷酸(Xu5P)。通过生化和结构分析发现,来自球形红杆菌Rhodobacter sphaeroides的CbbY蛋白(RsCbbY)及其在拟南芥中保守的同源物质AtCbbYA都是高度特异的XuBP磷酸酶。近日德国慕尼黑大学生物系Thilo Rühle课题组在Nature Communications上发表了题为“An ancient metabolite damage-repair system sustains photosynthesis in plants”的研究论文,证明了一种古老的代谢修复系统在植物的光合作用中起着重要作用。通过修复XuBP的磷酸化状态,植物能够更有效地利用二氧化碳进行光合作用,促进生长和发育。

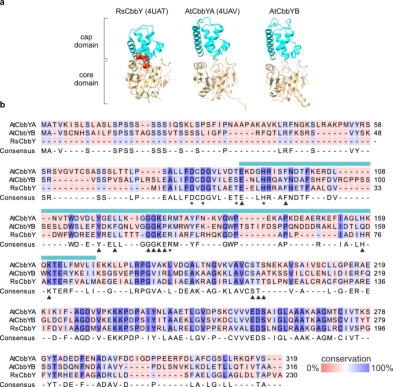
图1. CbbY蛋白的结构和序列比对
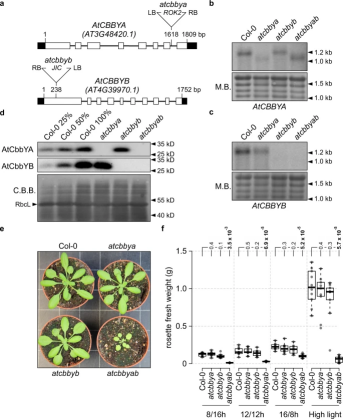
图2. atcbbyab双突变体拟南芥生长发育受阻
作者首先对拟南芥中与光合作用相关的蛋白质进行分析,针对拥有绿色细胞的光合生物共享的候选基因和光合作用相关的特异性基因进行了重点研究。在拟南芥中发现了两个HAD家族基因AtCbbYA和AtCbbYB,对关键催化位点和底物结合位点的保守性分析表明这些蛋白质在XuBP的去磷酸化过程中具有功能上的相似性(图1)。通过筛选,找到了AtCBBYA和AtCBBYB的Arabidopsis T-DNA插入突变系,并通过杂交获得了双突变体atcbbyab(图2a)。在atcbbya和atcbbyb突变体中没有观察到明显的表型,但与野生型植物相比,在所有测试条件下,atcbbyab双突变体的生长明显受到抑制(图2e,f)。这表明AtCbbYA和AtCbbYB在光合作用过程中的Rubisco损伤修复具有相同的功能。
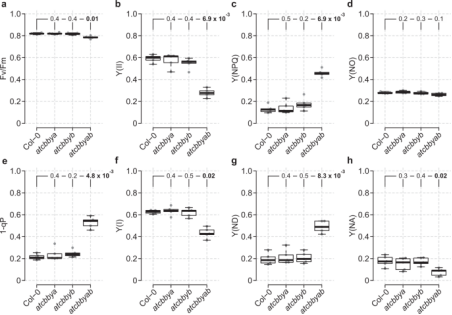 图3. AtCbbYA和AtCbbYB的缺失损害光合电子传递
图3. AtCbbYA和AtCbbYB的缺失损害光合电子传递

图4. 高效固碳依赖于AtCbbYA和AtCbbYB
为了量化AtCbbYA和AtCbbYB功能丧失对CBB循环活性的影响,作者进行了气体交换测量,并同时监测叶绿素a荧光(图4)。将Rubisco小亚基(RbcS)敲除突变体rbcs1a3b-1作为CBB循环活性降低的对照,发现缺乏AtCbbYA和AtCbbYB的情况下,AtCbbYA和AtCbbYB对于维持高效的光合电子传递和二氧化碳同化是必需的。与rbcs1a3b-1不同,atcbbyab的碳固定速率降低可能是由于在缺乏AtCbbYA和AtCbbYB的情况下对Rubisco抑制剂的降解效率不高。
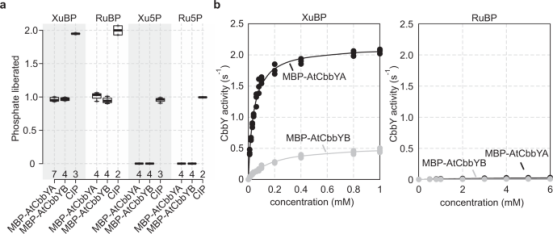 图5. MBP-ATCBBYA和MBP-ATCBBYB 特异性去磷酸化木糖酮-1,5-二磷酸酯 (XuBP)
研究人员通过在大肠杆菌中异源表达AtCbbYA和AtCbbYB并融合到麦芽糖结合蛋白(MBP)上,并利用淀粉麦芽糖基质进行纯化。结果显示,这两种融合蛋白可以分别将XuBP和RuBP分子释放出一个磷酸根,但无法去磷酸化木糖-5-磷酸(Xu5P)和核糖-5-磷酸(Ru5P)。相比之下,具有广泛底物特异性的小肠碱性磷酸酶(CIP)可以分别将XuBP或RuBP分子释放出两个磷酸根,并且也能够去磷酸化Xu5P和Ru5P。这些结果表明,AtCbbYB与AtCbbYA一样,可以特异地在XuBP和RuBP的1号位点切割磷酸根。
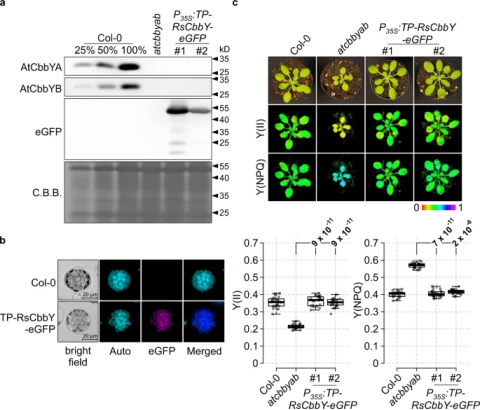
将Rhodobacter sphaeroides中发现的XuBP磷酸酶(RsCbbY)的CDS序列优化密码子后与预测的AtCbbYA质体转运肽的编码序列融合。将融合序列放置在花椰菜花叶病毒35S启动子的控制下,与eGFP编码的报告基因融合转入atcbbyab突变体中可以恢复拟南芥的AtCbbYA和AtCbbYB缺失表型(图6)。
图6. 当靶向叶绿体时,球形红杆菌(RsCbbY)的XuBP磷酸酶在拟南芥中起作用
图7. 通过CbbY磷酸酶调控CBB循环和光合作用的模型
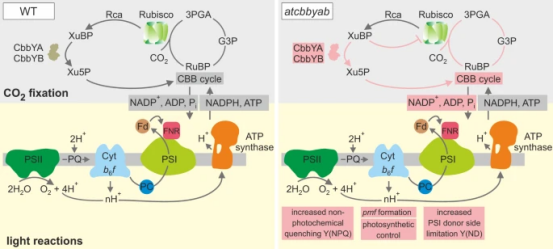
研究表明通过Rubisco活化酶介导的XuBP去磷酸化不足以保持高效光合作用,除非CbbYA/CbbYB降解系统可用于防止抑制剂的再结合。尽管碳同化速率有所降低,但atcbbyab突变体的表型并未导致致死(图2)。这表明在缺乏AtCbbYA和AtCbbYB的情况下,还存在另外一个过程来清除XuBP。该研究不仅揭示了植物中代谢物损伤修复系统的重要性,还对优化光合生物中的碳固定效率具有重要的影响。通过了解和操控这一代谢途径,我们可以寻找方法来提高植物的光合效率,从而增加农作物的产量为粮食安全和可持续农业的发展提供新的思路和方法。
图5. MBP-ATCBBYA和MBP-ATCBBYB 特异性去磷酸化木糖酮-1,5-二磷酸酯 (XuBP)
研究人员通过在大肠杆菌中异源表达AtCbbYA和AtCbbYB并融合到麦芽糖结合蛋白(MBP)上,并利用淀粉麦芽糖基质进行纯化。结果显示,这两种融合蛋白可以分别将XuBP和RuBP分子释放出一个磷酸根,但无法去磷酸化木糖-5-磷酸(Xu5P)和核糖-5-磷酸(Ru5P)。相比之下,具有广泛底物特异性的小肠碱性磷酸酶(CIP)可以分别将XuBP或RuBP分子释放出两个磷酸根,并且也能够去磷酸化Xu5P和Ru5P。这些结果表明,AtCbbYB与AtCbbYA一样,可以特异地在XuBP和RuBP的1号位点切割磷酸根。
将Rhodobacter sphaeroides中发现的XuBP磷酸酶(RsCbbY)的CDS序列优化密码子后与预测的AtCbbYA质体转运肽的编码序列融合。将融合序列放置在花椰菜花叶病毒35S启动子的控制下,与eGFP编码的报告基因融合转入atcbbyab突变体中可以恢复拟南芥的AtCbbYA和AtCbbYB缺失表型(图6)。
图6. 当靶向叶绿体时,球形红杆菌(RsCbbY)的XuBP磷酸酶在拟南芥中起作用
图7. 通过CbbY磷酸酶调控CBB循环和光合作用的模型
研究表明通过Rubisco活化酶介导的XuBP去磷酸化不足以保持高效光合作用,除非CbbYA/CbbYB降解系统可用于防止抑制剂的再结合。尽管碳同化速率有所降低,但atcbbyab突变体的表型并未导致致死(图2)。这表明在缺乏AtCbbYA和AtCbbYB的情况下,还存在另外一个过程来清除XuBP。该研究不仅揭示了植物中代谢物损伤修复系统的重要性,还对优化光合生物中的碳固定效率具有重要的影响。通过了解和操控这一代谢途径,我们可以寻找方法来提高植物的光合效率,从而增加农作物的产量为粮食安全和可持续农业的发展提供新的思路和方法。