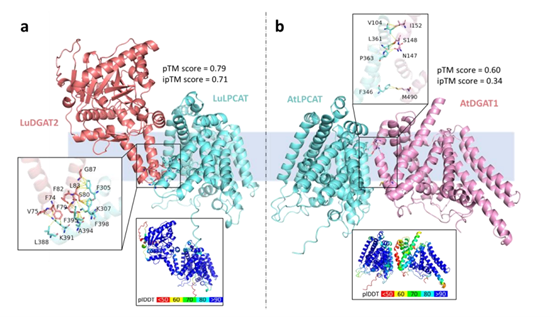
植物脂质通过细胞膜形成、能量储存和信号传递等功能,在植物发育和胁迫反应中发挥着重要的生物学作用。近年来,越来越多的证据表明,在脂质生物合成途径中的各种蛋白质参与者可以形成动态的蛋白质/蛋白质-脂质作用体或复合物,与植物代谢相互作用体/代谢物的新兴领域相呼应。此外,动态的脂质代谢作用体可能存在并在其他生物的脂质代谢中发挥重要作用,但我们目前对这些作用体/代谢物性质的了解还非常有限。2023年2月10日,Plant Biotechnology Journal杂志在线发表了加拿大阿尔伯塔大学的一篇题为“Protein interactomes for plant lipid biosynthesis and their biotechnological applications的综述论文。该文综述了参与植物脂质生物合成的蛋白质相互作用体的特征,重点强调了由不同亚基组成的蛋白质复合物以及由协同途径中的酶形成的瞬时或动态相互作用体;还展示了使用AlphaFold2程序预测了具有代表性的脂质生物合成相互作用体的结构,并讨论了在植物脂质生物技术中,使用相互作用体知识的现状以及未来的应用。由不同亚基组成的蛋白质复合体,包括乙酰辅酶a羧化酶(ACCase),脂肪酸合成酶(FAS)和脂肪酸延长酶(FAE)。高等植物中的乙酰辅酶a羧化酶(ACCase)有两种形式:由四个亚基组成的异源ACCase,形成多酶复合物,以及具有所有四种酶组分连接成单个多肽的同源ACCase。所有植物的胞浆中都有一个同源异构体,这有助于长链脂肪酸的生物合成和其他代谢过程。为了进一步了解植物中的异聚ACCase,本文利用深度学习结构预测程序AlphaFold2预测了拟南芥中α-CT/β-CT异四聚体的亚复合体结构。其中,拟南芥α-CT和β-CT的核心催化区与大肠杆菌α-CT、β-CT高度一致。其次,蛋白质-蛋白质相互作用对FAS的功能很重要,因为单独的II型FAS亚基,包括KAS、KAR、HAD和ENR,都是低聚物,并与ACP相互作用。来自大肠杆菌的II型FAS已被广泛研究,单个FAS亚基的结构已被解析,但这些亚基如何协同工作仍有待阐明。本文预测拟南芥KAR同四聚体(图1b)和HAD的结构时同源二聚体与两个ACP相互作用(图1c),使用AlphaFold2,预测的结构显示良好,与已知大肠杆菌直系物的结构模型能够很好地匹配。图一:使用AlphaFold2预测脂肪酸生物合成蛋白质复合物的结构。随后,对亚麻DGAT2与LPCAT、拟南芥DGAT1与LPCAT蛋白相互作用结构的预测结果表明,前者的预测可信度高于后者(图2),这可能是由于跨膜结构简单的膜酶(如DGAT2)比具有多个跨膜结构域的膜酶(如DGAT1)更容易建模。尽管如此,两个预测都表明了螺旋8、9和10LPCAT可能与DGAT1(图2b)和DGAT2(图2a)形成相互作用界面。在这些预测中确定了几个特定的相互作用位点,未来可以通过生化手段进一步验证。图2:利用AlphaFold2预测了亚麻LPCAT和DGAT2以及拟南芥LPCAT和DGAT1的蛋白-蛋白相互作用结构。总之,系统地研究脂质生物合成酶背景下相互作用体的形成过程是非常有意义的。这对于在我们在了解蛋白质相互作用体的基础上,设计新的基因工程方法上具有创新意义,并且对植物脂质生物技术的进一步发展至关重要。文章来源:植物生物技术Pbj