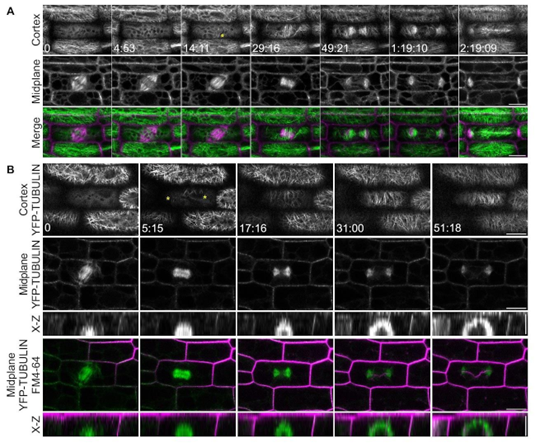
细胞分裂和正确的分裂方向对植物的发育和生长都很重要。细胞分裂是由细胞中部的一种称为胞膜的结构开始的。成膜细胞由包括微管在内的细丝组成,向外扩张形成新的细胞壁。膈肌定位是由定位在未来分裂位置或分裂位点的环中的蛋白质介导的。目前尚不清楚这些蛋白质如何促进分裂定位。

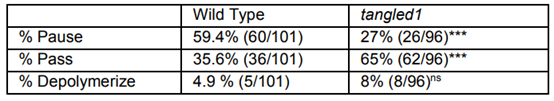
为了评估皮质末期微管与成纤维细胞微管相比是否具有不同的特性,用微管不稳定剂丙酰胺(5µM)处理玉米表皮细胞,并检查了与分裂位点相互作用的单个微管以及细胞内的其他位置。当皮质末期微管与分裂位点相互作用时,59%弯曲,表明微管在分裂位点短暂捕获时仍在生长。分裂位点的中间暂停或捕获时间为15秒,而其他位置为10.0秒。分裂位点微管加端的瞬时稳定可能促进整体垂直定向。
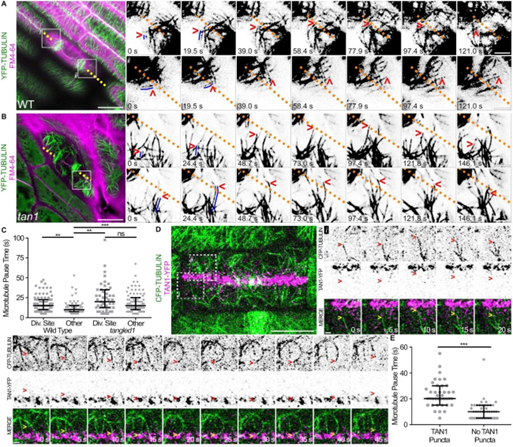
使用表达TAN1-YFP和CFP-TUBULIN的野生型细胞的延时成像测量了加末端的静止期微管。TAN1-YFP在分裂末期在分裂部位形成离散点,发现在野生型玉米细胞中,皮层终末期微管在TAN1-YFP点附近的停留时间比分裂位点内的其他区域长。检测了玉米TAN1突变体中的皮质末期微管。近30%的tan1细胞皮质终末期微管阵列稀疏或缺失。与野生型细胞相比,tan1突变细胞的皮层终末期微管在分裂位点没有瞬时稳定,与其他皮层位置相比,分裂位点的静止期微管没有显著差异。由于微管增加,加上分裂位点的末端停顿或捕获,皮质末期微管通常朝向分裂位点,并且皮质末期微管最常通过低接触角的平行捆绑添加到隔膜中。
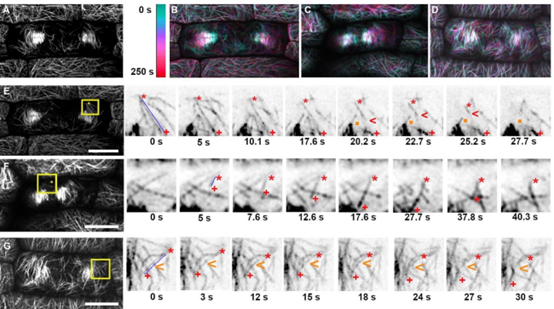
为了研究皮质末期微管阵列如何和成膜细胞相互作用,研究人员使用了延时成像。当皮质终末期微管与隔膜状体接触时,大多数(78%,n=197/252个微管,来自3种植物的5个细胞)通过平行束扎结合到隔膜状体的前缘。使用时间推移成像,测量了隔膜膜随时间的运动,并测量了相应的皮质末期微管阵列。将成膜细胞轨迹与分裂平面“上方”和“下方”皮质末期微管的相对积累进行了比较,结果表明皮质末期微管的持续积累,无论是在上方还是下方,都与成膜细胞在同一方向上的运动相关。与野生型植物相比,突变体成膜细胞扩张方向和皮层末期微管阵列积累的变化更大,但不对称的皮层末期微管积累与成膜细胞方向的变化之间的关系是相同的。
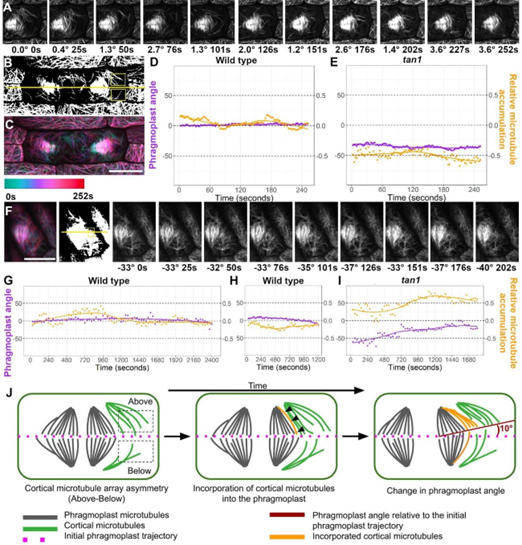
研究人员发现,在玉米的裂末期,在膜体到达细胞皮层之前,皮层的裂末微管阵列在玉米中积累并与分裂位点相互作用。虽然皮层末期微管的不对称积累和成膜细胞角度随时间的变化都可能独立于一些未知的线索,但笔者提出,通过平行束束结合到成膜细胞中的末期微管可以微调成膜细胞的位置,使其到达细胞皮层的精确分裂位点。由局部皮层微管提供的引导机制,当隔膜在细胞皮层膨胀时直接结合到隔膜中,将提供靶向所需的特异性。当成膜细胞与细胞皮层的距离在微米范围内时,将皮质末期微管直接加入成膜细胞以改变其局部位置。关于微管是否参与分裂定位,以及TANGLED1如何调节其在其他植物细胞中的动态,目前尚不清楚,有待于进一步研究。
文章来源:植物生物技术Pbj
