本文内容速览:
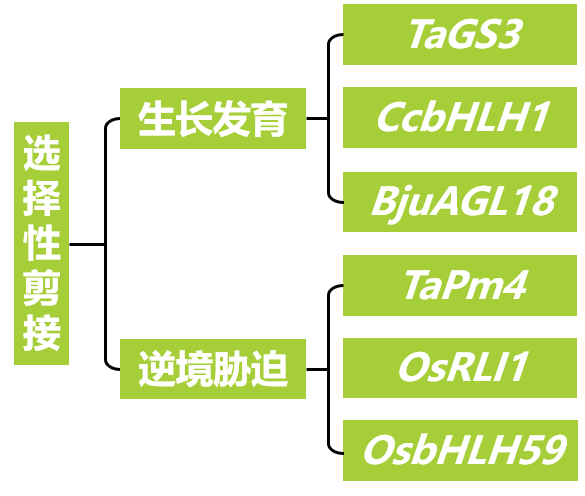
在“植物基因的“断舍离”——选择性剪接”一文中,小远给大家介绍了选择性剪接的基础知识,包括剪接的概念、剪接过程、常见的剪接类型以及选择性剪接的鉴定方法,如果对这部分知识还不太了解,强烈建议你去读一读这篇文章哦!当基因发生选择性剪接后,会产生多个不同的转录本,每个转录本都可能编码具有独特功能的蛋白质,进而在植物生长发育、应对环境胁迫等诸多生理过程中发挥关键作用。那么,在今天的文章中,小远主要想探讨一个关键问题——在实际研究过程中,我们该如何探究不同转录本的功能差异呢?别着急,跟着小远一起看看几个具体的实例吧!
选择性剪接在植物生长发育上的研究
2021年10月,河北师范大学李俊明课题组在International Journal of Molecular Sciences杂志上发表了一篇题为“Alternative splicing of TaGS3 differentially regulates grain weight and size in bread wheat”的研究论文,作者鉴定了Gγ亚基基因TaGS3的五种剪接转录本(TaGS3.1–TaGS3.5)在籽粒大小和粒重决定中发挥着不同的功能。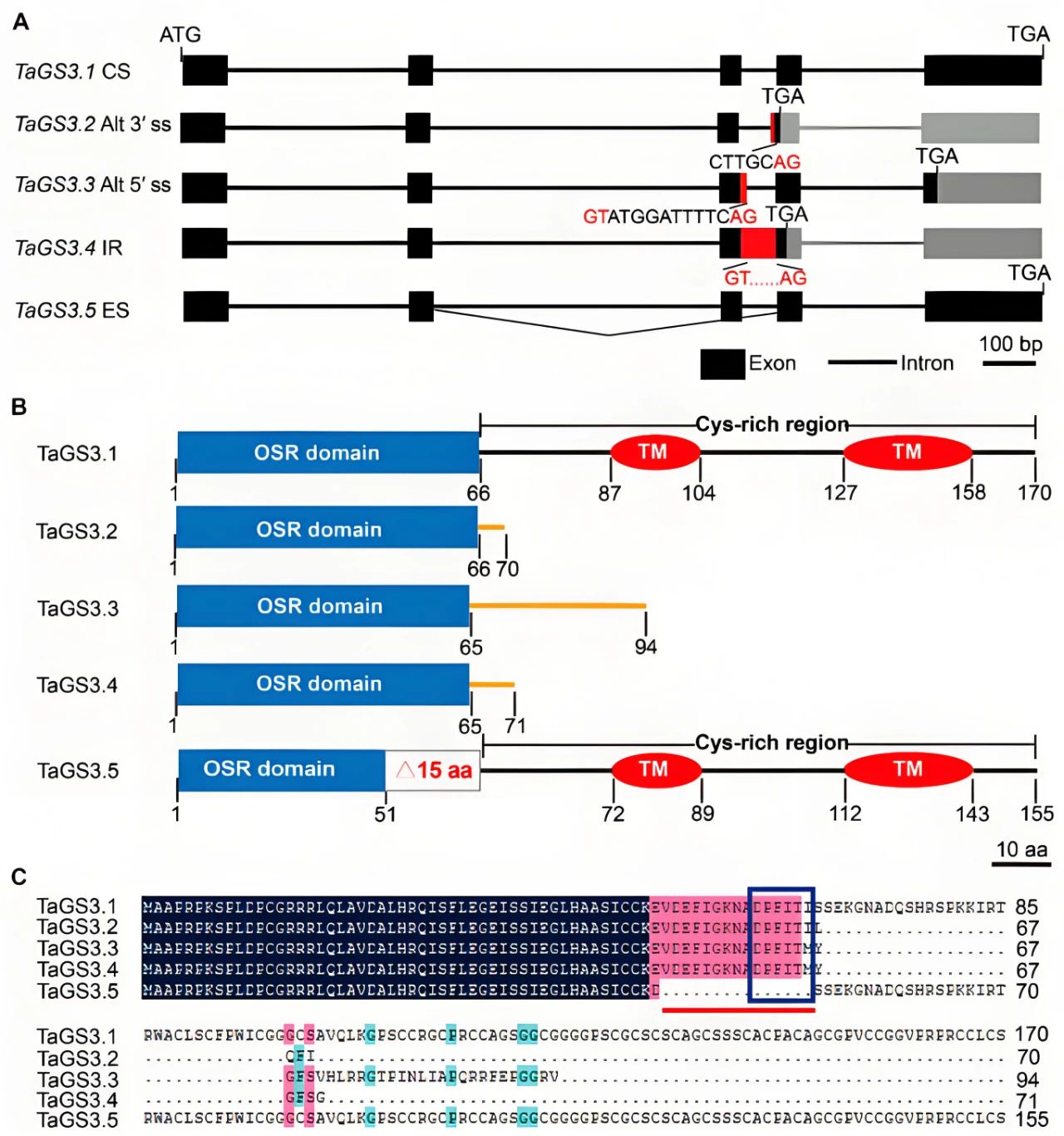 图1 TaGS3受到选择性剪接的影响(Ren et al., 2021)。(A)TaGS3剪接转录本的基因结构示意图;(B)五种TaGS3剪接转录本的蛋白结构示意图。TaGS3.1包含完整的OSR结构域(1-66aa)、两个跨膜结构域(87-104aa和127-158aa)和一个Cys富含区(67-170aa);TaGS3.2包含完整的OSR结构域,TaGS3.3和TaGS3.4中的OSR结构域有1aa的差异;TaGS3.5包含一个不完整的OSR结构域(1-51aa)和两个完整的跨膜结构域。OSR,器官大小调节结构域;TM,假定的跨膜结构域;橙色线表示C末端非Cys富含区;(C)TaGS3异构体的蛋白质序列比对。
图1 TaGS3受到选择性剪接的影响(Ren et al., 2021)。(A)TaGS3剪接转录本的基因结构示意图;(B)五种TaGS3剪接转录本的蛋白结构示意图。TaGS3.1包含完整的OSR结构域(1-66aa)、两个跨膜结构域(87-104aa和127-158aa)和一个Cys富含区(67-170aa);TaGS3.2包含完整的OSR结构域,TaGS3.3和TaGS3.4中的OSR结构域有1aa的差异;TaGS3.5包含一个不完整的OSR结构域(1-51aa)和两个完整的跨膜结构域。OSR,器官大小调节结构域;TM,假定的跨膜结构域;橙色线表示C末端非Cys富含区;(C)TaGS3异构体的蛋白质序列比对。
TaGS3.1、TaGS3.2、TaGS3.3和TaGS3.4在小麦不同发育时期的各种组织中均广泛表达,且表达模式相似,而在所有检测的组织中均未检测到TaGS3.5的表达(图2A)。TaGS3.1-GFP和TaGS3.5-GFP融合蛋白仅在质膜中观察到荧光,而TaGS3.2-GFP、TaGS3.3-GFP和TaGS3.4-GFP融合蛋白在质膜和细胞核中均可观察到荧光(图2B)。
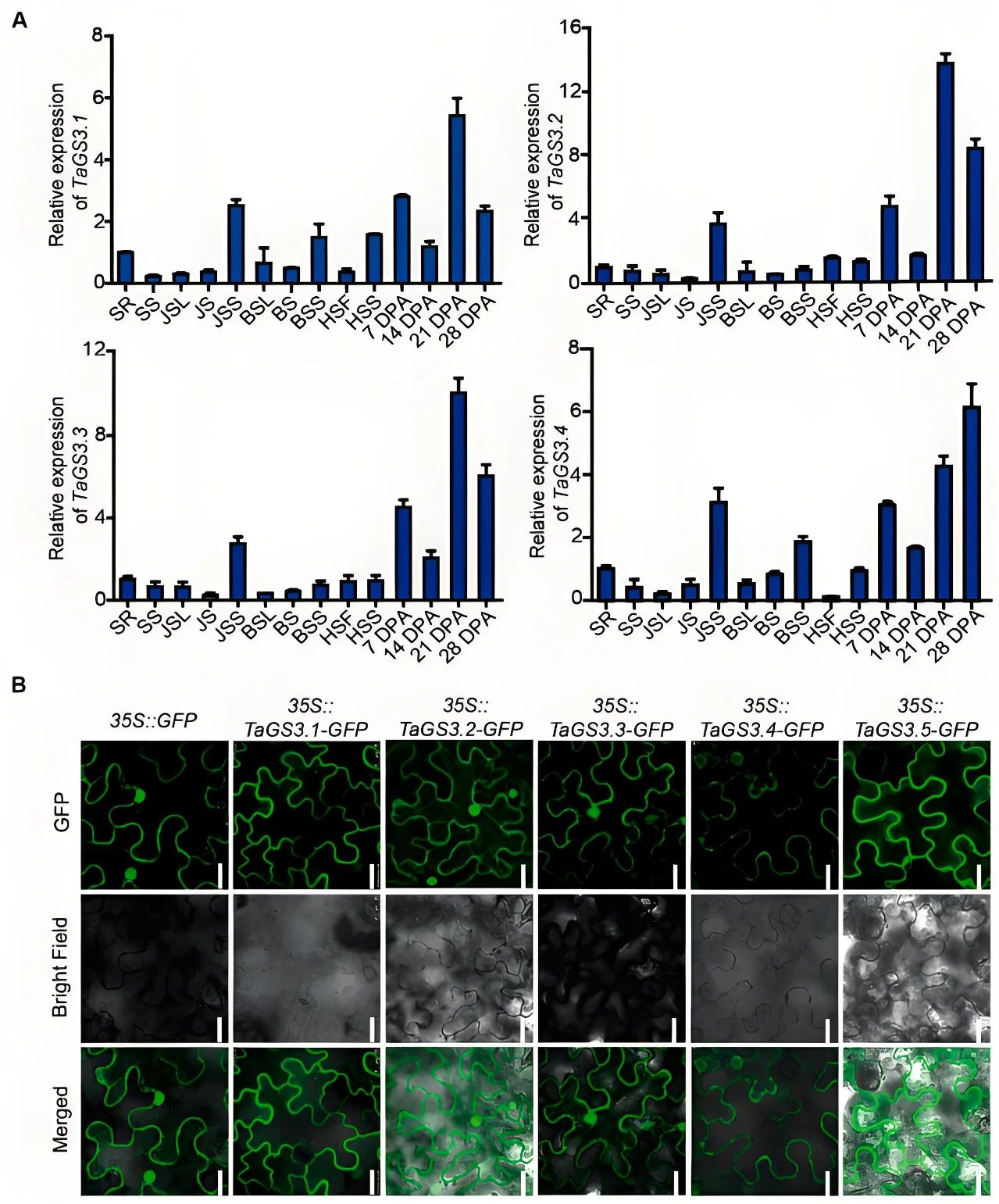
图2 TaGS3的表达模式和亚细胞定位(Ren et al., 2021)。(A)TaGS3的表达分析。SR,幼苗根;SS,幼苗芽;JSL,拔节期叶片;JS,拔节期茎;JSS,拔节期穗;BSL,孕穗期叶片;BS,孕穗期茎;BSS,孕穗期穗;HSF,抽穗期旗叶;HSS,抽穗期穗;7DPA-28DPA,开花后7-28天的籽粒;(B)TaGS3的亚细胞定位。
与野生型相比,TaGS3.1过表达株系显著降低了粒长、粒宽、千粒重和单株产量,而TaGS3.2过表达株系、TaGS3.3过表达株系和TaGS3.4过表达株系粒长、粒宽、千粒重和单株产量均没有显著差异,TaGS3.5过表达株系显著增加了粒长、粒宽、千粒重和单株产量(图3)。

图3 田间条件下TaGS3过表达株系与WT的性状表型比较(Ren et al., 2021)。(A)TaGS3剪接转录本过表达株系和WT整株表型;(B)TaGS3剪接转录本的T3转基因株系的免疫印迹分析;(C)转基因株系粒长(GL)比较;(D)转基因株系粒宽(GW)比较;(E)转基因株系千粒重(TGW)比较;(F)转基因株系单株产量(GY)比较。G蛋白由Gɑ、Gβ和Gγ亚基组成,Gβ介导的信号转导需要Gγ来形成活性Gβγ异二聚体。Y2H和Co-IP实验证实WGB1(Gβ)与TaGS3.1、TaGS3.2、TaGS3.3和TaGS3.4相互作用,但不与TaGS3.5相互作用,说明OSR结构域中的15个氨基酸残基对于TaGS3-WGB1相互作用的必要性,并且当C末端Cys富含区被删除时TaGS3与WGB1的互作会增强。酵母三杂实验表明TaGS3.2、TaGS3.3、TaGS3.4和TaGS3.1在与WGB1相互作用方面存在竞争。以上结果说明完整的OSR结构域是TaGS3作为粒长负调控因子发挥作用的必要条件,并且TaGS3中C末端Cys富集区与小麦籽粒大小呈负相关。
2024年11月,北京林业大学戴思兰课题组联合临沂大学邓成燕课题组在Ornamental Plant Research杂志上发表了一篇题为“Alternative splicing of activator CcbHLH1 gene accounts for anthocyanin absence in white cornflower”的研究论文,作者为揭示白色矢车菊(DTPW)中花青素缺失的分子机制,发现CcbHLH1在白色矢车菊花瓣中发生了选择性剪接,导致其长度缩短至1125bp,丧失了关键的螺旋-环-螺旋(helix-loop-helix, HLH)结构域以及全部ACT-like结构域。这一突变可能导致CcbHLH1功能丧失,因而被重新命名为CcbHLH1#(图4b)。CcbHLH1#在花瓣中特异表达,且在白色花瓣中的表达量显著高于在蓝色花瓣中的表达量(图4a)。CcbHLH1和CcbHLH1#均定位于细胞核中,选择性剪接并未改变其亚细胞定位(图4c)。Y2H和Split-LUC实验表明CcbHLH1和CcbHLH1#均能与CcMYB6-1发生相互作用(图5a、b)。Dual-LUC实验表明CcbHLH1#失去了激活CcDFR启动子转录活性的能力(图5c、d),这可能是白色矢车菊花瓣中花青素缺失的关键原因。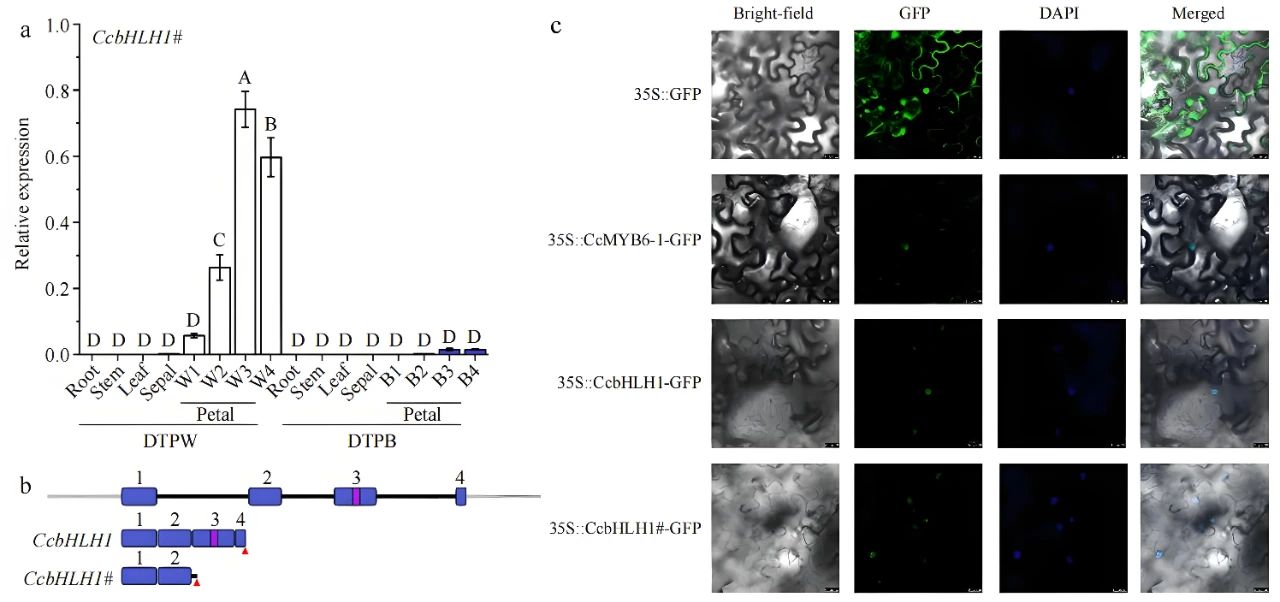 图4 CcbHLH1和CcbHLH1#的结构、表达模式和亚细胞定位(Deng et al., 2024)。(a)CcbHLH1#在DTPW(花瓣颜色为白色)和DTPB(花瓣颜色为蓝色)中的表达模式;(b)CcbHLH1和CcbHLH1#基因结构;(c)CcMYB6-1、CcbHLH1和CcbHLH1#的亚细胞定位分析。
图4 CcbHLH1和CcbHLH1#的结构、表达模式和亚细胞定位(Deng et al., 2024)。(a)CcbHLH1#在DTPW(花瓣颜色为白色)和DTPB(花瓣颜色为蓝色)中的表达模式;(b)CcbHLH1和CcbHLH1#基因结构;(c)CcMYB6-1、CcbHLH1和CcbHLH1#的亚细胞定位分析。
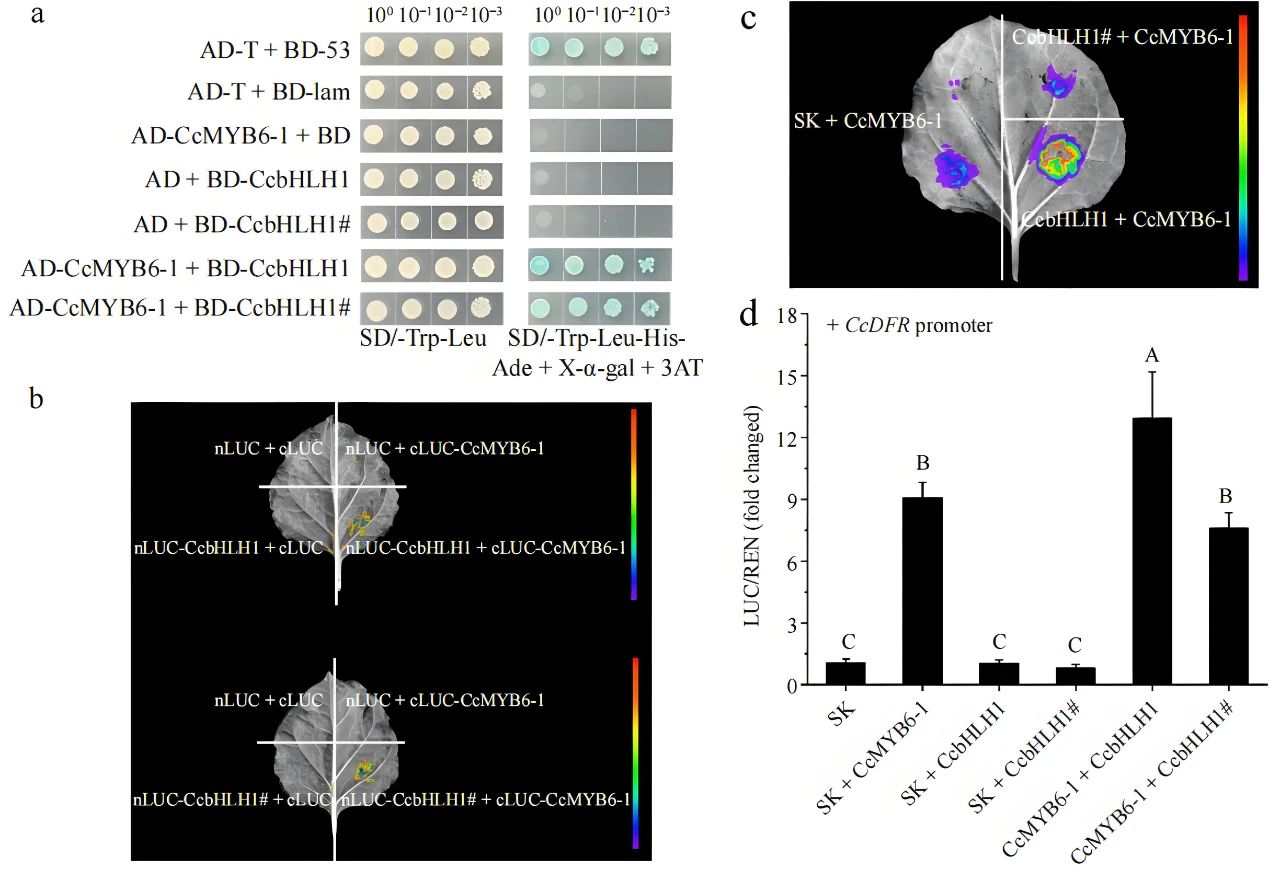
图5 CcbHLH1和CcbHLH1#调控花青素积累的分子机制解析(Deng et al., 2024)。(a)Y2H验证CcbHLH1和CcbHLH1#与CcMYB6-1的相互作用;(b)Split-LUC验证CcbHLH1和CcbHLH1#与CcMYB6-1的相互作用;(c、d)Dual-LUC验证CcbHLH1和CcbHLH1#结合CcDFR启动子的能力。
2025年3月,西南大学汤青林课题组在The Crop Journal杂志上发表了一篇题为“Modulation of flowering by an alternatively spliced AGL18-1 transcript in Brassica juncea”的研究论文,作者从芥菜中分离了BjuAGL18-1基因,并鉴定了其两个转录本:BjuAGL18-1L和BjuAGL18-1S,而且发现这两种转录本通过不同的调控机制参与芥菜光周期诱导的开花。BjuAGL18-1L和BjuAGL18-1S均定位在细胞核中(图6C)。此外,两者表达模式不同,BjuAGL18-1L和BjuAGL18-1S分别在短日照和长日照条件下被大量诱导(图6D、E)。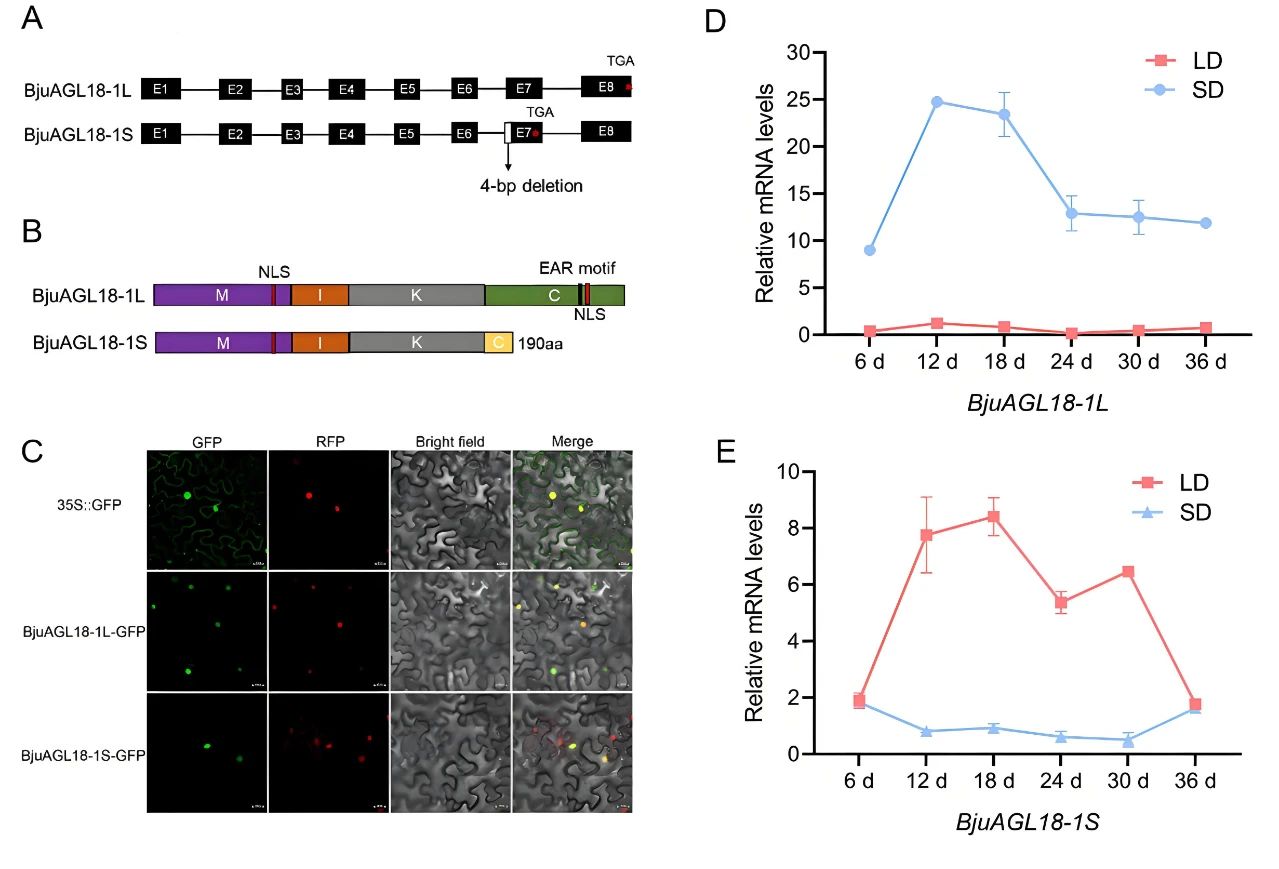 图6 BjuAGL18-1的序列分析、亚细胞定位和表达模式分析(Deng et al., 2025)。(A)BjuAGL18-1第六和第七外显子之间的内含子结构。与BjuAGL18-1L相比,BjuAGL18-1S在第七个外显子处有4bp的缺失引入了过早终止密码子,导致BjuAGL18-1S缺失EAR基序;(B)BjuAGL18-1的两种蛋白质异构体;(C)BjuAGL18-1L和BjuAGL18-1S蛋白的亚细胞定位;(D、E)qRT-PCR分析长日照(LD)和短日照(SD)处理的芥菜中的BjuAGL18-1L和BjuAGL18-1S的表达。
图6 BjuAGL18-1的序列分析、亚细胞定位和表达模式分析(Deng et al., 2025)。(A)BjuAGL18-1第六和第七外显子之间的内含子结构。与BjuAGL18-1L相比,BjuAGL18-1S在第七个外显子处有4bp的缺失引入了过早终止密码子,导致BjuAGL18-1S缺失EAR基序;(B)BjuAGL18-1的两种蛋白质异构体;(C)BjuAGL18-1L和BjuAGL18-1S蛋白的亚细胞定位;(D、E)qRT-PCR分析长日照(LD)和短日照(SD)处理的芥菜中的BjuAGL18-1L和BjuAGL18-1S的表达。
过表达BjuAGL18-1L显著延迟芥菜和拟南芥开花,而过表达BjuAGL18-1S则加速开花,进一步分析发现,BjuAGL18-1L可显著抑制开花相关基因BjuFUL和BjuFT的表达,而BjuAGL18-1S则促进开花相关基因BjuFUL和BjuFT的表达(图7)。其具体情况是:Y2H、BiFC、Split-LUC以及Dual-LUC实验发现,BjuAGL18-1L通过EAR基序与BjuAFR2、BjuHDA9形成蛋白复合体,并协同BjuAGL15直接结合在开花基因BjuFUL的启动子区域,以抑制该基因表达,从而延缓开花。而BjuAGL18-1S因缺失EAR基序无法形成抑制复合体,反而通过异源二聚化干扰BjuAGL18-1L功能,削弱其对下游基因的抑制作用,从而促进开花。
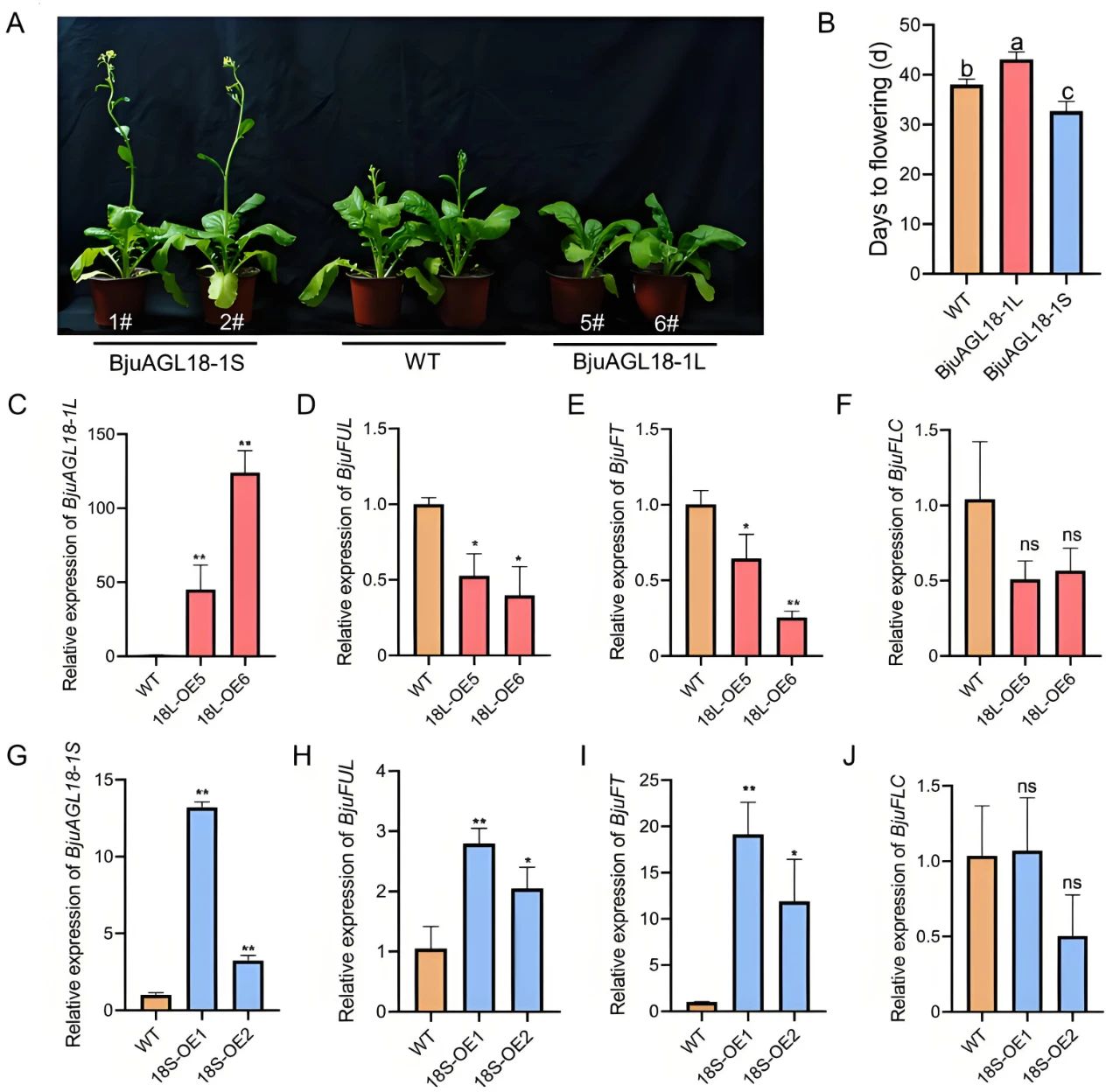
图7 BjuAGL18-1过表达导致芥菜出现相反的开花表型(Deng et al., 2025)。(A、B)与野生型(WT)株系相比,BjuAGL18-1L过表达转基因株系的开花时间明显延迟,而BjuAGL18-1S过表达转基因株系的开花时间明显早于WT株系;(C-F)BjuAGL18-1L-OE转基因株系中BjuAGL18-1L和开花相关基因BjuFT、BjuFUL和BjuFLC的表达量;(G-J)BjuAGL18-1S-OE转基因株系中BjuAGL18-1S和开花相关基因BjuFT、BjuFUL和BjuFLC的表达量。
2021年3月,苏黎世大学Beat Keller课题组在Nature Plants杂志上发表了一篇题为“Wheat Pm4 resistance to powdery mildew is controlled by alternative splice variants encoding chimeric proteins”的研究论文,作者克隆并深入研究了小麦白粉病小种特异性抗病基因Pm4b,该基因由七个外显子组成,其中外显子6和外显子7以互斥方式进行选择性剪接,产生两个剪接转录本Pm4b_V1和Pm4b_V2。感染白粉病后Pm4_V1和Pm4_V2的表达水平没有显著差异,表明Pm4b_V1和Pm4b_V2 对抗病性的贡献相似。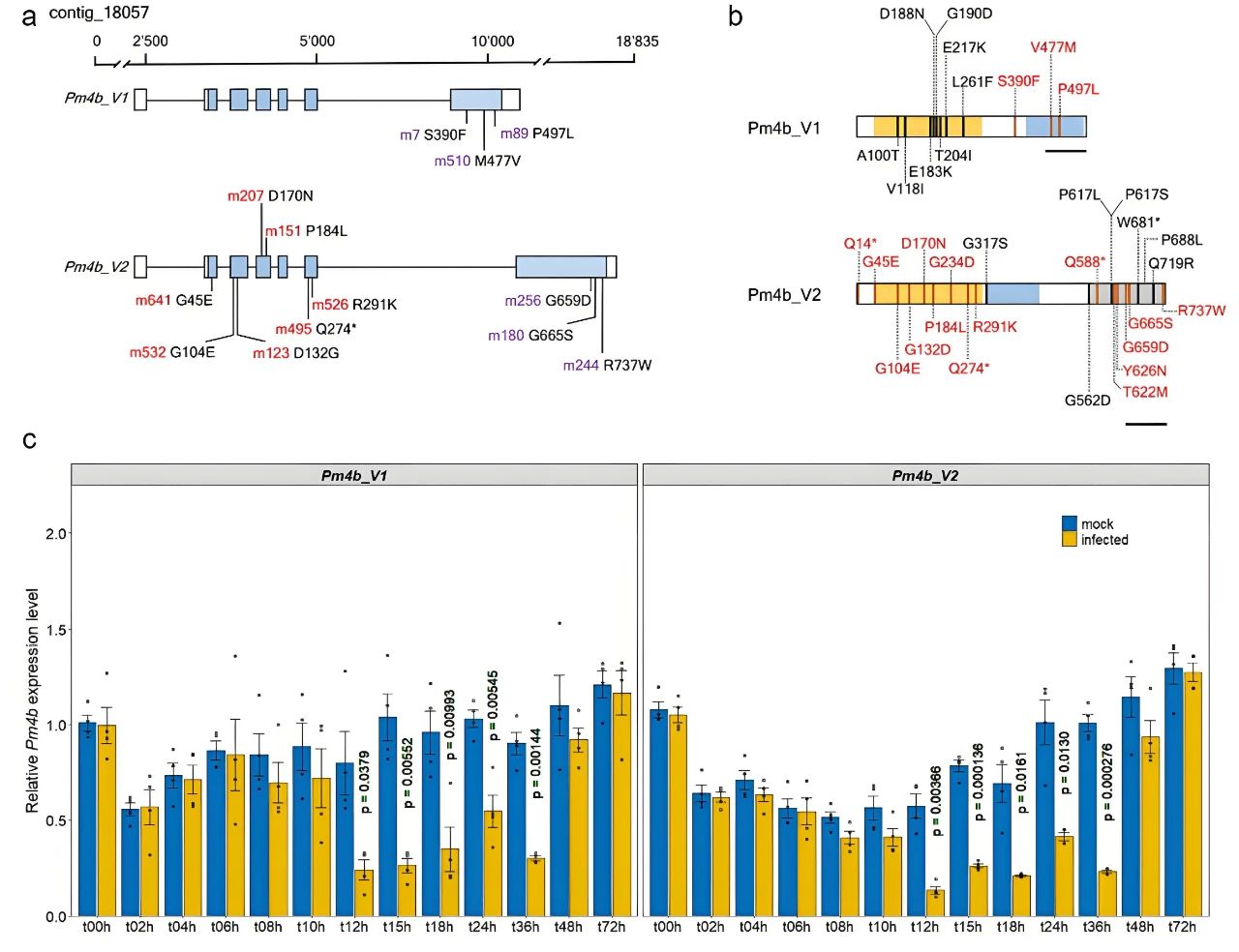 图8 Pm4b的分子鉴定和表达分析(Sánchez-Martín et al., 2021)。(a)Pm4b的基因结构和可变剪接;(b)Pm4b_V1和Pm4b_V2蛋白质异构体;(e)接种Bgt96224的Fed-Pm4b植株中Pm4_V1和Pm4_V2剪接转录本的转录水平。
图8 Pm4b的分子鉴定和表达分析(Sánchez-Martín et al., 2021)。(a)Pm4b的基因结构和可变剪接;(b)Pm4b_V1和Pm4b_V2蛋白质异构体;(e)接种Bgt96224的Fed-Pm4b植株中Pm4_V1和Pm4_V2剪接转录本的转录水平。
作者将含有Pm4b_V1和Pm4b_V2两个全长cDNA的基因转入对Bgt96224菌株敏感的小麦品种‘Bobwhite S26’中,发现Pm4b基因的两个剪接转录本Pm4b_V1和Pm4b_V2都是赋予小麦对白粉病抗性所必需的。同时通过VIGS技术进一步测试Pm4b介导的对小麦白粉病的抗性,发现Pm4b_V1或Pm4b_V2的沉默导致小麦对白粉病的抗性降低(图9)。
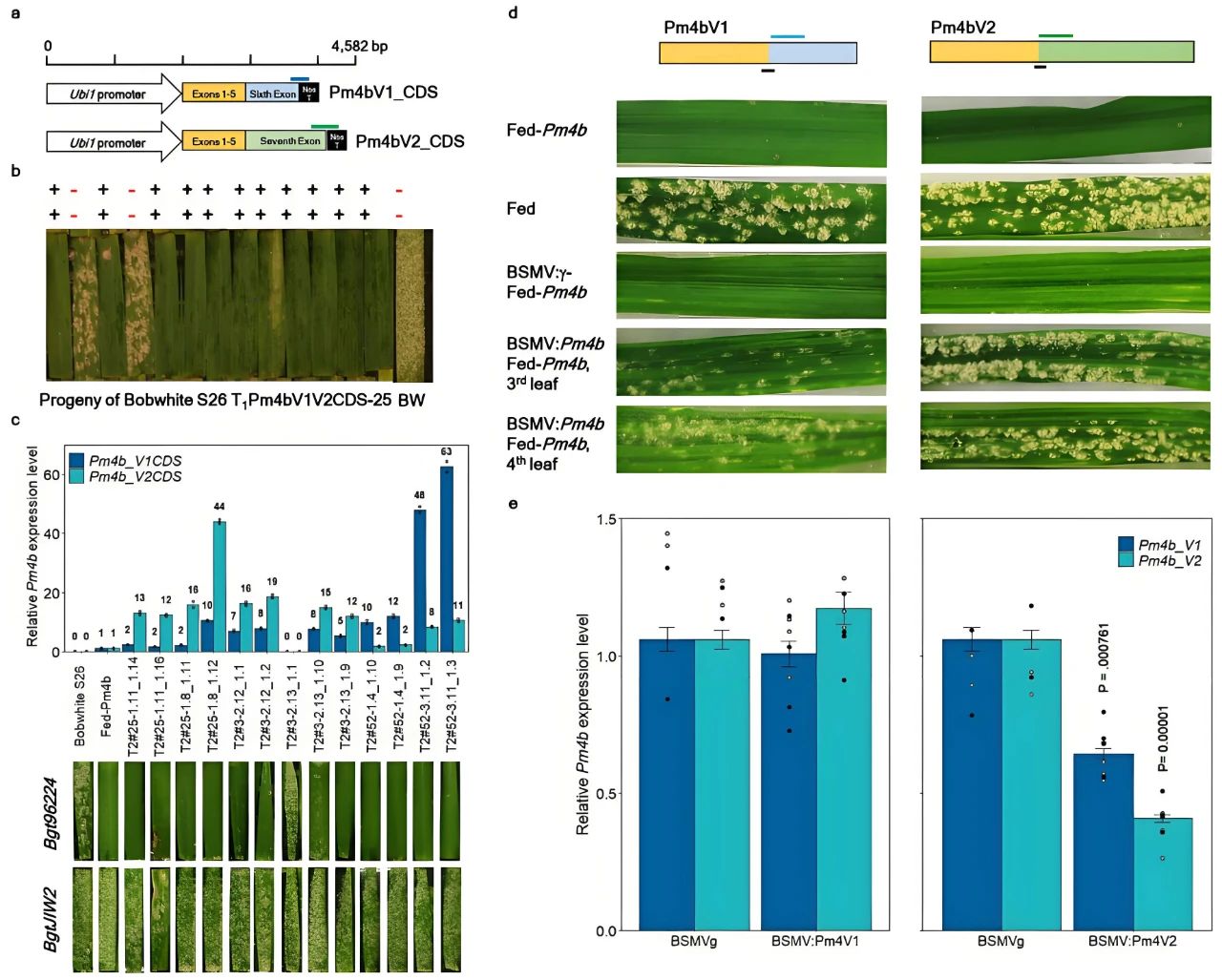
图9 通过转基因互补和VIGS技术验证Pm4b基因的功能(Sánchez-Martín et al., 2021)。(a)用于转化易感Bobwhite S26(BW)的两种构建体示意图;(b)对来自T1 Pm4bV1V2CDS-25的T1代后代进行筛选。Pm4bV1_CDS(上排)和Pm4bV2_CDS(下排)转基因的存在(+)或缺失(-)情况与所测试的各个T1代植株的抗病/感病表型相对应;(c)转基因T2代株系中Pm4bV1_CDS和Pm4bV2_CDS转基因的表达水平以及抗病性检测;(d)经过VIGS处理并在用对Pm4b无毒性的Bgt96224分离株感染后,代表性植株第三片和第四片叶子的症状;(e)通过qRT-PCR评估BSMVg、BSMV:Pm4V1和BSMV:Pm4V2感染的Fed-Pm4b植株的Pm4bV1和Pm4bV2的表达水平。
最后,作者结合一系列分子实验,解析了Pm4介导抗病性的可能分子机制(图10)。在白粉病菌侵染植物的过程中,当病菌附着在植物表面并侵入寄主细胞后,会形成吸器,在吸器形成的早期阶段(侵染后12h-24h)会释放低水平的AvrPm4效应子,此时小麦细胞已经通过Pm4b基因介导的机制开始抵抗病原体的入侵。在吸器形成阶段(侵染后48h),会大量释放AvrPm4效应子,从而诱导更强的由Pm4介导的防御反应,进而产生超敏反应(HR)。在没有受到病原菌侵染时,Pm4_V1和Pm4_V2处于静息状态,形成一个通过C2域相互作用的异源复合体,这个复合体锚定在内质网上,并且是未激活的(图10)。当AvrPm4被Pm4的C2C/D或激酶域识别后,异源复合体发生构象变化,导致激酶活性的激活(图10),并启动抗病性。
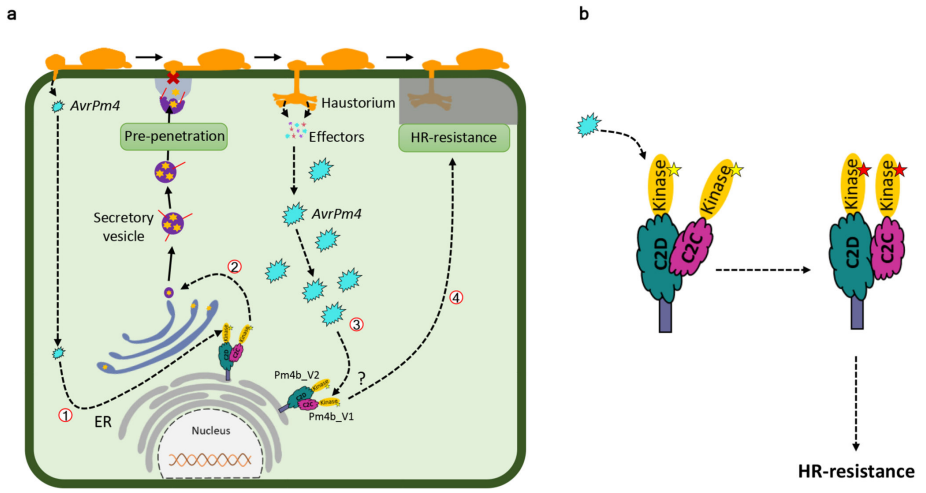
图10 Pm4介导小麦抗病性的可能工作模型(Sánchez-Martín et al., 2021)。
2022年5月,中国农业科学研究院农业资源与农业区划研究所易可可/阮文渊课题组在The Plant Cell杂志上发表了一篇题为“Alternative splicing of REGULATOR OF LEAF INCLINATION 1 modulates phosphate starvation signaling and growth in plants”的研究论文,作者发现REGULATOR OF LEAF INCLINATION 1(RLI1)基因的可变剪接可调节磷酸盐信号转导和植物在磷饥饿胁迫下的生长。
作者发现叶倾角调节因子RLI1的可变剪接在水稻中产生两种蛋白亚型:第五内含子的保留导致RLI1a中出现提前终止密码子,从而编码MYB转录因子,而RLI1b中第五内含子的剪接使其能够编码MYB-CC蛋白(图11)。
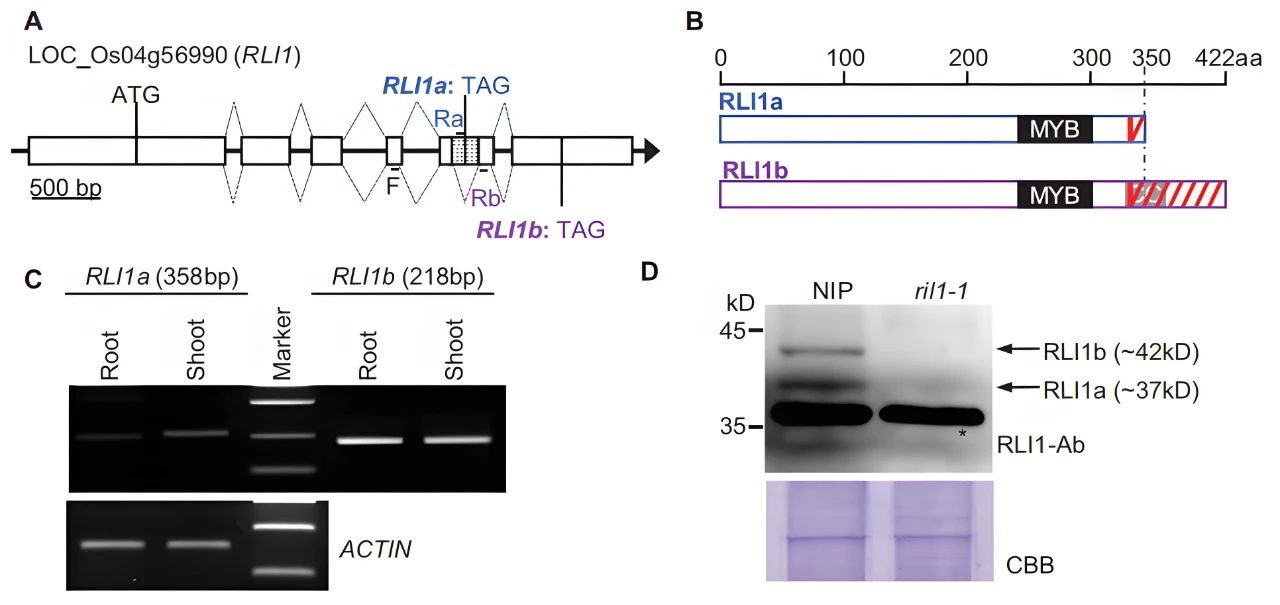
图11 RLI1产生两个转录本及其相应的蛋白质亚型(Guo et al., 2022)。(A)RLI1的基因结构;(B)RLI1a和RLI1b的蛋白结构;(C)RLI1不同转录本亚型的RT-PCR分析;(D)RLI1a和RLI1b蛋白质亚型的免疫印迹分析。
接着作者构建了RLI1a过表达株系和RLI1b过表达株系,发现RLI1a-Ox的叶片倾角显著大于WT,但RLI1b-Ox的叶片倾角与WT相似(图12A-D)。同时测定RLI1a-Ox和RLI1b-Ox植株的叶片关节细胞长度,发现RLI1a-Ox的细胞长度明显大于WT,而RLI1b-Ox的细胞长度和WT之间没有显著差异(图12E、F),进一步证明RLI1a通过影响叶片关节细胞伸长来调节叶片倾斜度,而RLI1b则不会。后续实验中作者通过ChIP-qPCR和Dual-LUC实验证明RLI1a通过调节油菜素类固醇的生物合成以影响叶片倾斜度。
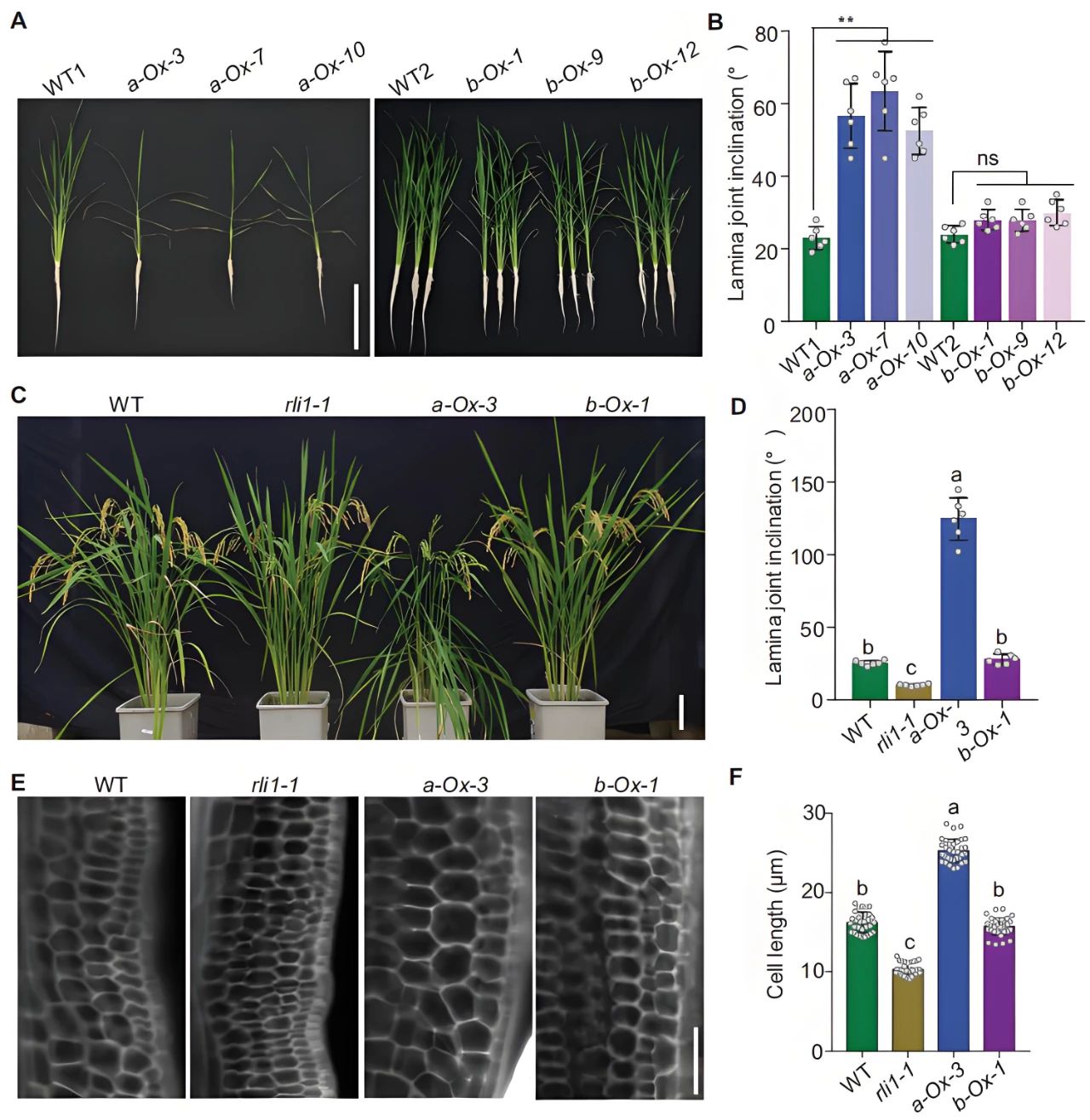
图12 RLI1a而非RLI1b调节叶片倾斜度(Guo et al., 2022)。(A、B)水培条件下WT、RLI1a-Ox和RLI1b-Ox的表型分析(A)和叶片倾斜度分析(B),使用主分蘖顶端第三片叶子分析叶片倾斜度;(C、D)土壤中生长的WT、rli1突变体、RLI1a-Ox和RLI1b-Ox的表型分析(C)和叶片倾斜度分析(D),使用从顶部数第二片叶子测量叶片倾斜度;(E)WT、rli1突变体、RLI1a-Ox和RLI1b-Ox第二片顶部叶子的叶片关节纵向切片。
接着作者发现在磷(Pi)充足的情况下RLI1a-Ox株系和RLI1b-Ox株系均表现出典型的叶尖坏死症状,并且RLI1a-Ox和RLI1b-Ox植株的茎部Pi积累均高于WT。而在Pi缺乏的条件下,RLI1a-Ox株系和RLI1b-Ox株系叶尖坏死症状消失(图13A-C)。同时作者发现,RLI1a-Ox和RLI1b-Ox植株的Pi饥饿信号和Pi吸收相关基因IPS1、SPX1、VPE2以及Pi转运基因PT2和PT10均被显著诱导(图13D)。结果表明,RLI1a和RLI1b均在调节水稻Pi积累和Pi饥饿信号中发挥作用。
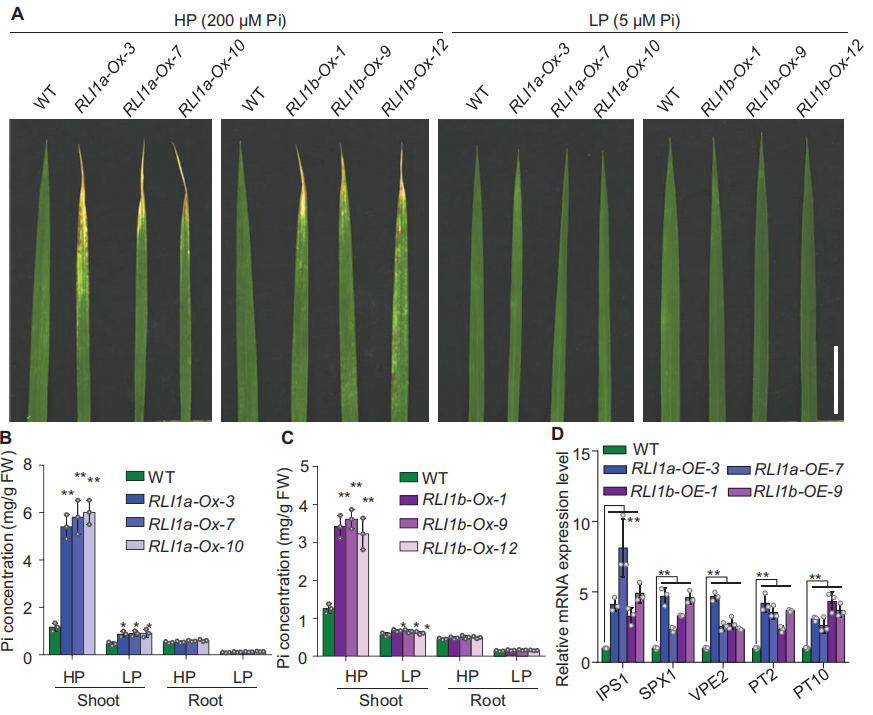
图13 RLI1a和RLI1b均调节Pi积累和Pi饥饿信号(Guo et al., 2022)。(A)WT、RLI1a过表达株系和RLI1b过表达株系叶片的表型。植物在Pi充足(HP,200μM Pi)和缺乏(LP,5μM Pi)条件下生长30天。使用主分蘖顶端第三片叶子进行分析;(B、C)在HP和LP条件下生长的WT、RLI1a-Ox和RLI1b-Ox的Pi测量值;(D)WT、RLI1a-Ox和RLI1b-Ox植株中IPS1、SPX1、VPE2、PT2和PT10的相对表达水平。
此外作者结合一系列分子实验,提出了RLI1的选择性剪接和RLI1蛋白质稳定性如何与PHR2协同调节Pi信号转导和植物生长,以增强对Pi缺乏胁迫的适应性的工作模型(图14)。在Pi充足条件下,RLI1a和RLI1b转录水平相似,但RLI1b和PHR2的蛋白质稳定性较低,而RLI1a的蛋白质稳定性较高,且对P1BS基序的亲和力较低,导致PSI基因的中度激活并促进油菜素内酯(BL)生物合成和信号转导。这些过程有助于植物在Pi充足条件下有效维持适合生长的Pi和BR含量。在Pi缺乏条件下,RLI1b转录水平较高,而RLI1a转录水平较低,RLI1b和PHR2的稳定性升高导致PSI基因的激活,从而促进Pi的吸收和利用。此外,在Pi缺乏条件下,RLI1a转录水平的降低和RLI1a蛋白质稳定性的降低会减弱BL生物合成和信号转导相关基因的激活,从而诱导形成直立枝条结构。这种直立枝条结构有助于植物减少Pi的消耗。
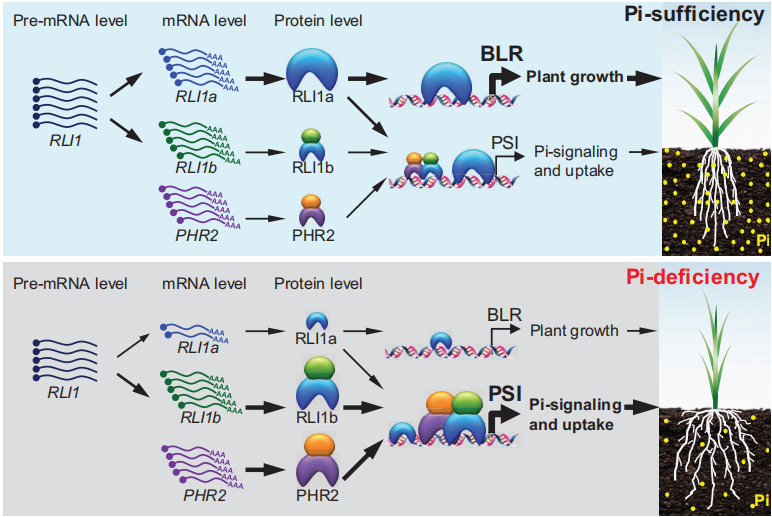
图14 RLI1在调节植物生长和Pi信号传导以帮助植物适应Pi缺乏的工作模型(Guo et al., 2022)。线条粗细表示影响程度。不同大小的蛋白质图表示蛋白质水平。RLI1a作为单体靶向下游基因。RLI1b和PHR2作为二聚体靶向下游基因。
2025年1月,云南大学黄立钰课题组、胡凤益课题组联合中国农业科学院作物科学研究所张帆课题组在Science Bulletin杂志上发表了一篇题为“Alternative splicing drives the functional diversification of a bHLH transcription factor in the control of growth and drought resistance in rice”的研究论文,揭示了OsbHLH59转录因子通过选择性剪接在调控水稻生长发育和抗旱性中发挥作用。
作者鉴定了两种不同的OsbHLH59转录本:OsbHLH59.1和OsbHLH59.2(图15a),它们的组织表达模式以及在干旱胁迫下的表达模式不同,此外,在干旱、PEG和ABA处理下,OsbHLH59.2的丰度与OsbHLH59.1的丰度的比率上升,表明OsbHLH59中发生了胁迫依赖性选择性剪接(图15b-i)。OsbHLH59.1和OsbHLH59.2两种蛋白质均可与G-box和E-box序列结合(图15j、k)。OsbHLH59.1和OsbHLH59.2都具有转录激活活性,其中OsbHLH49.2表现出更强的活性(图15m)。作者进一步证实OsbHLH59.1和OsbHLH59.2可以形成同源二聚体,并在植物细胞的细胞核内相互作用形成异二聚体(图15n)。然而,异二聚体表现出抑制的转录活性(图15l、m),表明OsbHLH59.1和OsbHLH59.2在转录调控水平上是彼此的竞争性阻遏物。
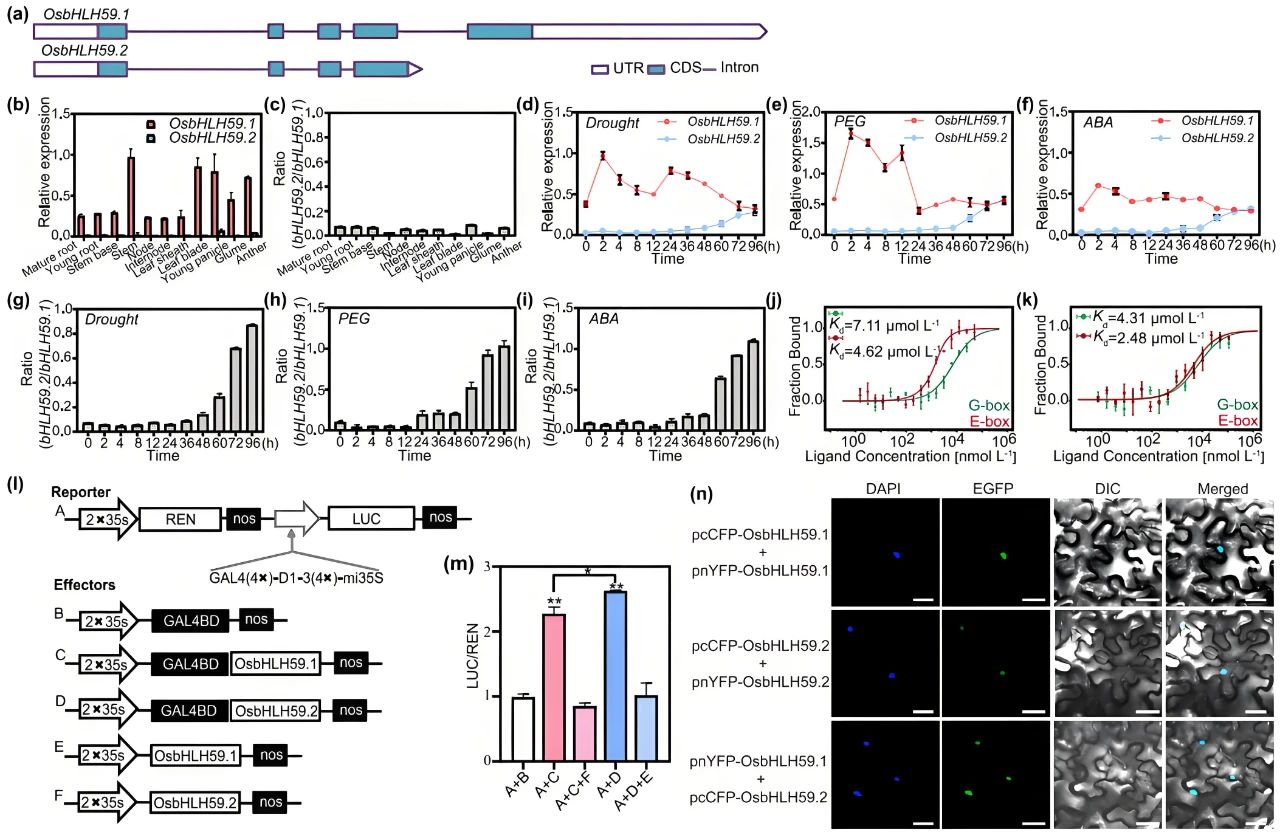 图15 OsbHLH59.1和OsbHLH59.2基因的结构和生化特性(Ning et al., 2025)。(a)OsbHLH59.1和OsbHLH59.2基因组序列图;(b、c)抽穗期野生型(WT)植株不同组织中的OsbHLH59转录本积累和OsbHLH59.2/OsbHLH59.1比率;(d-f)OsbHLH59.1和OsbHLH59.2在干旱、20%PEG和ABA处理下WT幼苗的表达水平分析;(g-i)响应环境刺激的OsbHLH59.2/OsbHLH59.1比率;(j、k)通过微量热泳动(MST)量化MBP-OsbHLH59.1和MBP-OsbHLH59.2与G-box、E-box的结合亲和力;(l、m)OsbHLH59.1和OsbHLH59.2的转录激活分析;(n)BiFC测定OsbHLH59.1和OsbHLH59.2在本氏烟草叶片细胞的细胞核中自身形成同型二聚体和异型二聚体。
与野生型相比,OsbHLH59.1-OE株系提高了株高,OsbHLH59.2-OE株系和OsbHLH59基因敲除株系株高降低(图16a)。OsbHLH59.1-OE株系中观察到的植株株高增加主要是由于细胞数量增加,而在OsbHLH59.2-OE和OsbHLH59-KO株系中观察到细胞数量减少(图16a-c)。OsbHLH59.1在bhlh59突变体中的过表达成功地恢复了bhlh59-8的生长抑制表型。然而,在bhlh59突变体中过表达OsbHLH59.2并没有挽救表型(图16d-f)。表明OsbHLH59.1是OsbHLH59的主要转录本,并在调节植物生长发育中发挥关键作用。
此外,作者评估了OsbHLH59.1和OsbHLH59.2对干旱胁迫的响应功能,干旱处理3d后,OsbHLH59.1-OE和OsbHLH59.2-OE株系的卷曲叶片均少于WT(图16g、h)。干旱处理5d后,水稻叶片全部卷曲,在复水7d后,OsbHLH59.1-OE和OsbHLH59.2-OE株系的幼苗存活率均显著高于WT,并且OsbHLH59.2-OE株系的存活率高于OsbHLH59.1-OE株系(图16g-i)。表明OsbHLH59.1和OsbHLH59.2均正向调控水稻的耐旱性,其中OsbHLH59.2表现出更强的作用。
图15 OsbHLH59.1和OsbHLH59.2基因的结构和生化特性(Ning et al., 2025)。(a)OsbHLH59.1和OsbHLH59.2基因组序列图;(b、c)抽穗期野生型(WT)植株不同组织中的OsbHLH59转录本积累和OsbHLH59.2/OsbHLH59.1比率;(d-f)OsbHLH59.1和OsbHLH59.2在干旱、20%PEG和ABA处理下WT幼苗的表达水平分析;(g-i)响应环境刺激的OsbHLH59.2/OsbHLH59.1比率;(j、k)通过微量热泳动(MST)量化MBP-OsbHLH59.1和MBP-OsbHLH59.2与G-box、E-box的结合亲和力;(l、m)OsbHLH59.1和OsbHLH59.2的转录激活分析;(n)BiFC测定OsbHLH59.1和OsbHLH59.2在本氏烟草叶片细胞的细胞核中自身形成同型二聚体和异型二聚体。
与野生型相比,OsbHLH59.1-OE株系提高了株高,OsbHLH59.2-OE株系和OsbHLH59基因敲除株系株高降低(图16a)。OsbHLH59.1-OE株系中观察到的植株株高增加主要是由于细胞数量增加,而在OsbHLH59.2-OE和OsbHLH59-KO株系中观察到细胞数量减少(图16a-c)。OsbHLH59.1在bhlh59突变体中的过表达成功地恢复了bhlh59-8的生长抑制表型。然而,在bhlh59突变体中过表达OsbHLH59.2并没有挽救表型(图16d-f)。表明OsbHLH59.1是OsbHLH59的主要转录本,并在调节植物生长发育中发挥关键作用。
此外,作者评估了OsbHLH59.1和OsbHLH59.2对干旱胁迫的响应功能,干旱处理3d后,OsbHLH59.1-OE和OsbHLH59.2-OE株系的卷曲叶片均少于WT(图16g、h)。干旱处理5d后,水稻叶片全部卷曲,在复水7d后,OsbHLH59.1-OE和OsbHLH59.2-OE株系的幼苗存活率均显著高于WT,并且OsbHLH59.2-OE株系的存活率高于OsbHLH59.1-OE株系(图16g-i)。表明OsbHLH59.1和OsbHLH59.2均正向调控水稻的耐旱性,其中OsbHLH59.2表现出更强的作用。
 图16 选择性剪接使OsbHLH59在控制生长和抗旱性方面的功能多样化(Ning et al., 2025)。(a)3周龄WT、bhlh59、OsbHLH59.1-OE和OsbHLH59.2-OE幼苗的表型;(b)通过石蜡切片对转基因植株幼苗期叶鞘进行细胞学观察;(c)WT、bhlh59、OsbHLH59.1-OE和OsbHLH59.2-OE植株的细胞数分析;(d-f)遗传互补系的表型(d)、株高(e)和根长统计(f);(g、h)转基因植株经干旱处理的幼苗表现;(i)复水后7天的幼苗存活率;(j)OsbHLH59介导的水稻植物生长与耐旱性之间权衡的工作模型。
最后作者通过一系列实验解析了OsbHLH59介导的水稻植物生长与耐旱性之间权衡的分子机制。在正常条件下,组成性表达的OsbHLH59.1通过激活关键靶基因正向调控水稻的生长发育。然而,在干旱胁迫下,OsbHLH59的选择性剪接被触发,导致通过ABA依赖性途径产生OsbHLH59.2。OsbHLH59.2与OsbHLH59.1相互作用形成异二聚体,抑制水稻的生长发育。随后,OsbHLH59.2激活靶基因,包括OsNCED4、OsMSR15、OsDREB1F和SNAC1,从而增强水稻的抗旱性(图16j)。
根据以上文章,我们来总结一下可以从哪些方面来探究植物基因不同转录本的功能差异。
图16 选择性剪接使OsbHLH59在控制生长和抗旱性方面的功能多样化(Ning et al., 2025)。(a)3周龄WT、bhlh59、OsbHLH59.1-OE和OsbHLH59.2-OE幼苗的表型;(b)通过石蜡切片对转基因植株幼苗期叶鞘进行细胞学观察;(c)WT、bhlh59、OsbHLH59.1-OE和OsbHLH59.2-OE植株的细胞数分析;(d-f)遗传互补系的表型(d)、株高(e)和根长统计(f);(g、h)转基因植株经干旱处理的幼苗表现;(i)复水后7天的幼苗存活率;(j)OsbHLH59介导的水稻植物生长与耐旱性之间权衡的工作模型。
最后作者通过一系列实验解析了OsbHLH59介导的水稻植物生长与耐旱性之间权衡的分子机制。在正常条件下,组成性表达的OsbHLH59.1通过激活关键靶基因正向调控水稻的生长发育。然而,在干旱胁迫下,OsbHLH59的选择性剪接被触发,导致通过ABA依赖性途径产生OsbHLH59.2。OsbHLH59.2与OsbHLH59.1相互作用形成异二聚体,抑制水稻的生长发育。随后,OsbHLH59.2激活靶基因,包括OsNCED4、OsMSR15、OsDREB1F和SNAC1,从而增强水稻的抗旱性(图16j)。
根据以上文章,我们来总结一下可以从哪些方面来探究植物基因不同转录本的功能差异。
1、转录本结构差异解析:分析不同转录本在基因结构层面的差异,包含外显子-内含子组成、剪接位点变化等;同时,对其编码蛋白的结构进行探究,包括蛋白的一级序列特征、二级和三级空间构象等,明确结构差异为后续功能研究奠定基础。

2、表达模式差异探究:分析转录本在植物不同组织(如根、茎、叶、花和果实等)以及不同生长发育阶段(从种子萌发到衰老)的表达变化规律;关注基因诱导表达差异,探究在生物胁迫(如病原菌侵染)或非生物胁迫(如干旱、高温、低温等)条件下转录本的表达响应;此外,明确转录本的亚细胞定位差异,确定其在细胞内的具体分布位置。

3、功能差异验证:通过构建不同转录本的过表达、沉默及敲除植株,对比观察不同株系的表型变化。

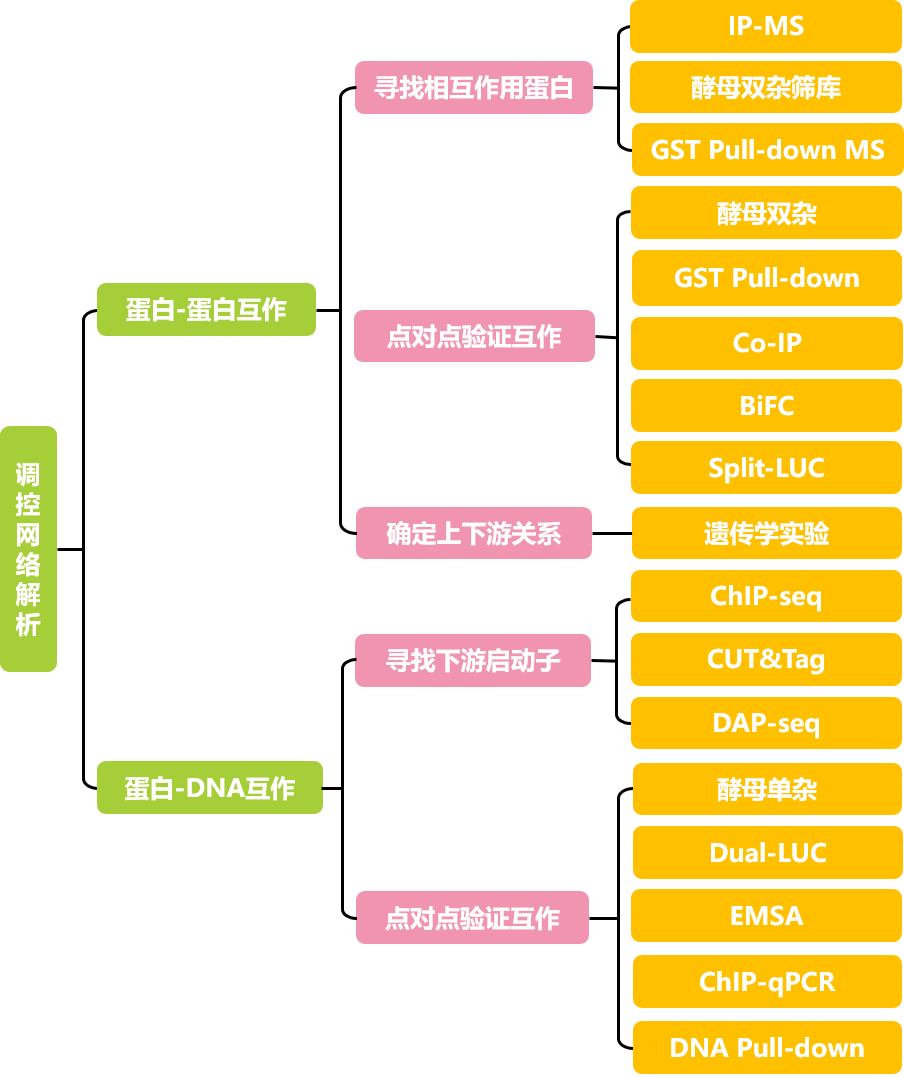
4、分子机制解析:借助蛋白-蛋白、蛋白-DNA互作验证实验,深入分析不同转录本调控网络的差异,从而揭示其存在功能差异的分子机制。
在这篇文章里,小远从植物生长发育与逆境胁迫两大维度出发,为大家列举了不同基因的选择性剪接转录本存在功能差异的参考文献,同时,还总结了探究植物基因不同转录本功能差异的研究思路,从剖析转录本结构差异,到探究表达模式差异,再到验证功能差异以及解析分子机制。希望小远的这些总结,能为大家的研究工作开拓新的思路,在探索植物基因奥秘的征程中发挥积极作用!
References:
Deng C, Zheng X, Wang J, et al. Alternative splicing of activator CcbHLH1 gene accounts for anthocyanin absence in white cornflower[J]. Ornamental Plant Research, 2024, 4(1).
Deng Q, Lu H, Liu D, et al. Modulation of flowering by an alternatively spliced AGL18-1 transcript in Brassica juncea[J]. The Crop Journal, 2025.
Guo M, Zhang Y, Jia X, et al. Alternative splicing of REGULATOR OF LEAF INCLINATION 1 modulates phosphate starvation signaling and growth in plants[J]. The Plant Cell, 2022, 34(9): 3319-3338.
Ning M, Li Q, Wang Y, et al. Alternative splicing drives the functional diversification of a bHLH transcription factor in the control of growth and drought tolerance in rice[J]. Science bulletin, 2025, 70(2): 153-156.
Ren X, Zhi L, Liu L, et al. Alternative splicing of TaGS3 differentially regulates grain weight and size in bread wheat[J]. International journal of molecular sciences, 2021, 22(21): 11692.
Sánchez-Martín J, Widrig V, Herren G, et al. Wheat Pm4 resistance to powdery mildew is controlled by alternative splice variants encoding chimeric proteins[J]. Nature Plants, 2021, 7(3): 327-341.
