本文内容速览:
在植物基因功能研究中,我们往往首先想到的是基因的过表达和敲除,即通过调控蛋白的“上调”(up-regulation)或“下调”(down-regulation)来探索其功能。然而,蛋白功能的研究是否仅仅局限于表达水平的调控?早在20世纪中期,Christian Anfinsen通过开创性的实验发现,蛋白的氨基酸序列决定了其三维结构,而这种结构又与蛋白的功能密切相关。由此可见,蛋白功能的研究不仅应关注表达水平的变化,还应包括对蛋白结构的设计与改造。
过去由于技术的限制以及已知蛋白结构数据的匮乏,植物领域在蛋白结构设计和改造方面进展缓慢,但近年来随着基因编辑技术的成熟、人工智能的飞速发展、蛋白工程工具的完善以及高通量筛选方法的普及,这一领域开始展现出巨大的潜力。在对关键功能蛋白进行优化、增强环境适应性以及开发新功能方面,蛋白结构设计与改造正逐渐显示出不可替代的价值。未来,在蛋白设计与改造的助力下,通过跨学科的深度合作与技术的全面整合,植物研究有望迈向更加精准和高效探索的新阶段。接下来,小远将给大家介绍植物蛋白功能研究为什么需要进行结构改造,再结合具体的文献案例讲述如何进行蛋白结构改造。
在植物分子生物学研究中,基因过表达和敲除通过改变基因的表达水平来探究其功能,从而为优化植物表型提供可能性。然而,简单地上调或下调某个基因的表达并不总能满足实际需求。一个典型的例子是,当过表达某些抗病相关蛋白时,虽然植物的抗病性得到了提高,但生长往往会受到限制,这一点在我们往期推文“植物的生长-防御平衡”中已有详细讨论,感兴趣的读者可以回顾相关内容。不仅是抗病与生长的矛盾,其他抗逆性状(如抗旱性)与生长的平衡问题同样难以兼顾。所以说,植物的确面临着“鱼与熊掌不可兼得”的窘境,而这一难题迫切需要一种新的方法来突破。蛋白结构改造正是能够解决这一问题的关键手段。简单来说,基因敲除和过表达主要作用于蛋白质的“量”的改变,而蛋白结构改造则通过调整蛋白本身的功能属性,带来“质”的提升。正是这种能力,使蛋白结构改造成为解决复杂性状平衡问题的理想工具。
蛋白质改造的一个经典例子是绿色荧光蛋白(GFP),它最早于1988年在维多利亚多管水母(Aequorea victoria)中被发现。自1994年起,GFP被首次应用于线虫神经元标记,并因其在发光成像中的独特性能,逐渐成为各种荧光标记实验中的核心工具。随着科学研究需求的不断变化,特别是对更稳定且多样化的荧光蛋白的需求增加,研究人员对原始的avGFP进行了一系列改造。最终,这些改造衍生出了如增强型绿色荧光蛋白(EGFP)、增强型蓝色荧光蛋白(EBFP)、增强型青色荧光蛋白(ECFP)和增强型黄色荧光蛋白(EYFP)等多种变体。这一过程展示了通过改变几个氨基酸序列,就能显著改变GFP的光学特性,甚至引发“质”的飞跃。
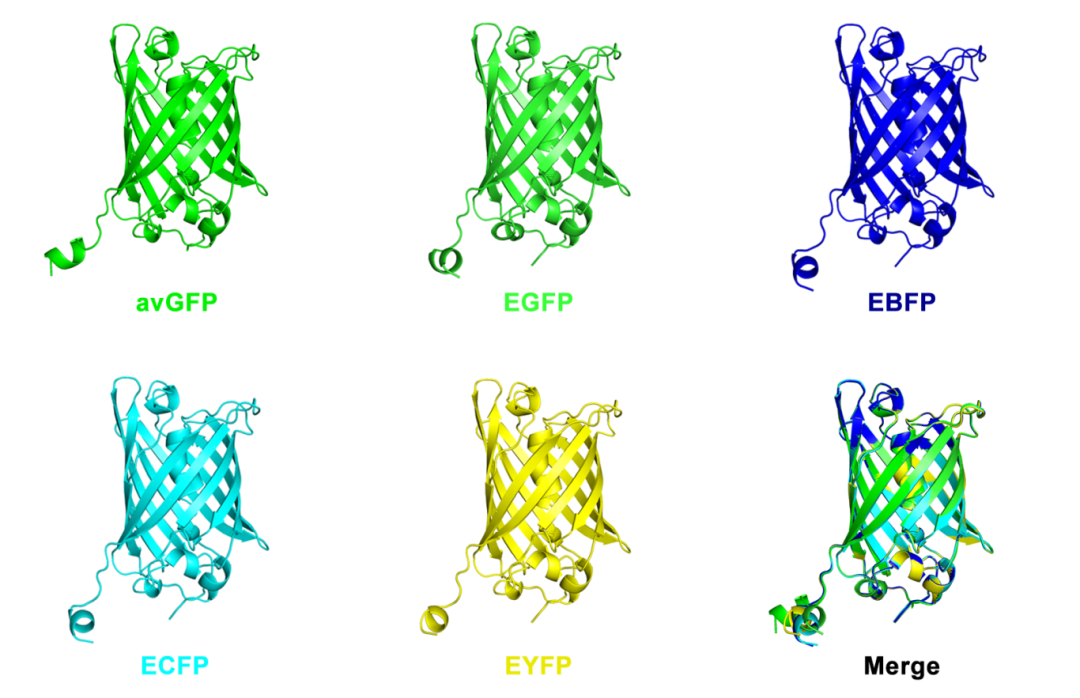
图1 原始avGFP及不同变体的结构。注:avGFP共有238个氨基酸。相比于avGFP,其余变体仅有几个位点氨基酸的不同。
前文提到,蛋白结构改造可以实现蛋白本身功能属性的改变,那么在植物研究的过程中具体该如何进行蛋白结构改造呢?小远总结了一下,大概有以下几种方法,分别是直接突变法、智能设计法、高通量筛选法、功能调控辅助法以及随机突变法。
直接突变法
直接突变是一种基础且广泛应用的蛋白结构改造手段,它通过对蛋白的结构直接引入突变来实现功能优化。在实施直接突变之前,首先需要明确哪些氨基酸位点的突变可能导致蛋白结构或功能的显著变化。通过深入分析这些关键位点,可以精准定位潜在的改造目标。随后,结合先进的基因编辑工具,对目标蛋白的关键氨基酸残基进行定点突变,从而实现对蛋白功能的精确调控和优化。
2024年9月,中国农业大学朱旺升课题组在Molecular Plant杂志上发表了一篇题为“Natural variations of maize ZmLecRK1 determine its interaction with ZmBAK1 and resistance patterns to multiple pathogens”的研究论文。实验前期,作者通过GWAS鉴定出一个抗病蛋白ZmLecRK1(G型凝集素受体激酶)。作者发现特定的ZmLecRK1等位基因可赋予玉米对多种病原菌的抗性,通过实验发现ZmLecRK1结构域中天然存在的A404变体提高了ZmLecRK1-ZmBAK1互作以及ZmLecRK1相关蛋白复合物的形成,从而引起下游免疫信号的激活,这也是含有A404变体材料具有更强抗病性的关键(图2A、C、D)。在高粱和水稻的ZmLecRK1蛋白中,通过基因编辑将404位的S替换为A可大大增强其诱导烟草细胞坏死的能力(图2B)。
图2 含A404的LecRK1是植物抗病所必需的(Li et al., 2024)。(A)抗性材料中ZmLecRK1Qi319 A404突变为S后引起烟草坏死反应严重减弱,易感材料中ZmLecRK1TY4 S404突变为A后引起烟草坏死反应增强;(B)烟草中的LecRK1和水稻中的LecRK6将相同的位点S突变为A后,引起烟草坏死反应显著增强;(C)通过AlphaFold2-Multimer预测了ZmLecRK1Qi319 A404变成S后与ZmBAK1的互作强度减弱;(D)Co-IP进一步证明了A404是ZmLecRK1Qi319与ZmBAK1互作的关键。
智能设计依托现代结构生物学与人工智能技术,逐渐成为蛋白改造的核心策略。通过结构生物学解析或者AI工具的辅助,可以预测蛋白质结构在突变后的可能构象变化。这不仅加速了功能优化的设计过程,还提供了定向进化的候选突变区域。AlphaFold 3这一工具在植物蛋白结构改造的应用在往期推文“诺奖级工具AlphaFold在研究中的应用”进行了详细的介绍,感兴趣的读者可以回顾相关内容。
2024年11月,中国农业大学彭友良课题组在JIPB杂志上发表了一篇题为“A resurfaced sensor NLR confers new recognition specificity to non-MAX effectors”的研究论文。水稻存在一类重要的成对NLR免疫受体RGA4和RGA5,RGA5利用金属离子结合结构域(heavy metal-associated,HMA)特异识别稻瘟菌中的效应蛋白AVR-Pia和AVR1-CO39,并激活RGA4介导的抗病反应。基于这种独特的识别机制,作者想人为设计新的RGA5去识别稻瘟菌中的效应蛋白。首先,作者以效应蛋白AVR-Pita作为靶点,筛选了与之互作的HMA蛋白,然后将筛选出来的HMA蛋白的HMA结构域与RGA5的HMA结构域进行互换,实验发现改造后的RGA5HMA120能够识别AVR-Pita并激活RGA4引起的细胞死亡(图3)。此外,表达RGA5HMA120的转基因水稻并不会在产量等重要农艺性状上有明显的改变。
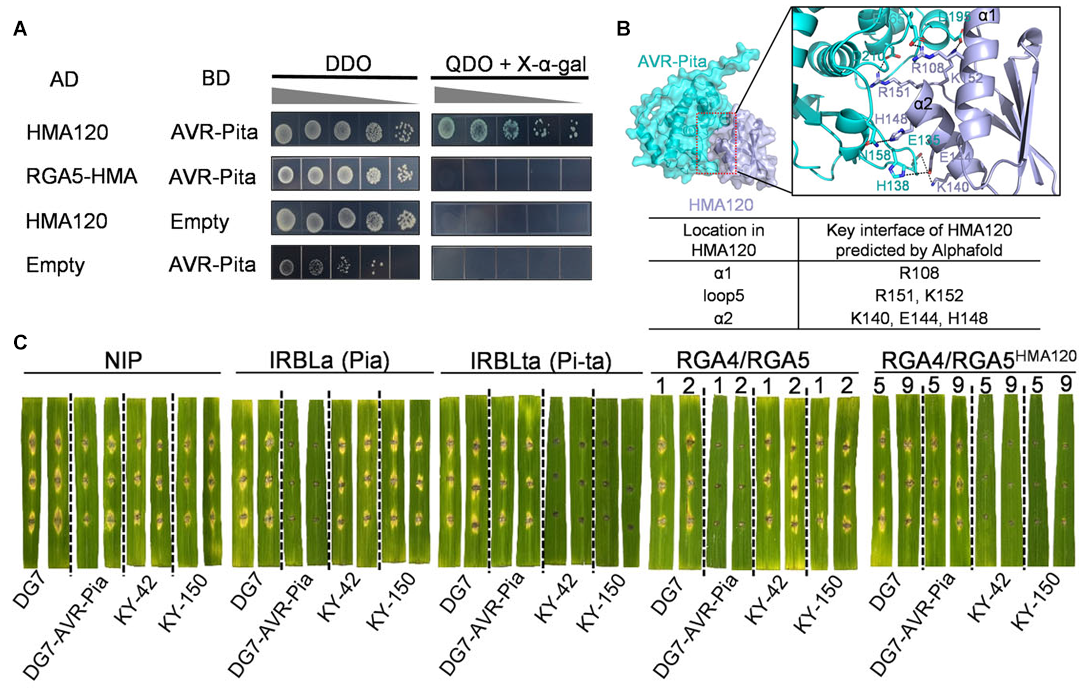
图3 新改造的RGA5HMA120能够识别AVR-Pita并引起免疫反应(Zhu et al., 2024)。(A)通过酵母筛选与AVR-Pita互作的HMA;(B)通过AlphaFold2预测AVR-Pita与HMA120的互作界面;(C)过表达RGA4/RGA5HMA120的水稻材料对携带AVR-Pita的稻瘟菌菌株KY-42和KY-150表现出抗性。
高通量筛选主要包括定向进化与高通量筛选两部分。尽管在前文提到,自然条件下的有利变异可以直接用于蛋白改造,但由于自然变异速率较低,可利用的有利变异相对有限。但借助基因编辑这一强有力的工具,可以快速产生大量突变体。随后,通过高效的筛选,有望获得符合要求的蛋白结构变异,为功能优化和改造提供更多可能性。
2024年2月,华中农业大学金双侠课题组和石河子大学聂新辉课题组联合在Genome Biology杂志上发表了一篇题为“Precise fine-turning of GhTFL1 by base editing tools defines ideal cotton plant architecture”的研究论文。植物成花素(FT)和抗成花素(TFL1)通过调控营养生长和生殖生长,对植物的株型结构起到重要的决定作用。然而,敲除GhTFL1会导致棉花植株出现极端矮化和极早开花的现象,这严重限制了其育种价值。为此,作者尝试对GhTFL1进行高密度碱基突变,从而生成大量突变体,期望筛选出株高适中、果枝缩短、株型紧凑、生育期缩短的棉花新种质。通过筛选一系列突变体,作者成功发现了GhTFL1L86P突变体(图4)。该突变体表现出果枝缩短、开花提前以及顶花表型的特征,且顶花与腋花均聚集在主茎上(图4B)。这一突变体在一定程度上克服了GhTFL1敲除突变体无法用于育种的局限性,为棉花育种提供了更为理想的选择。
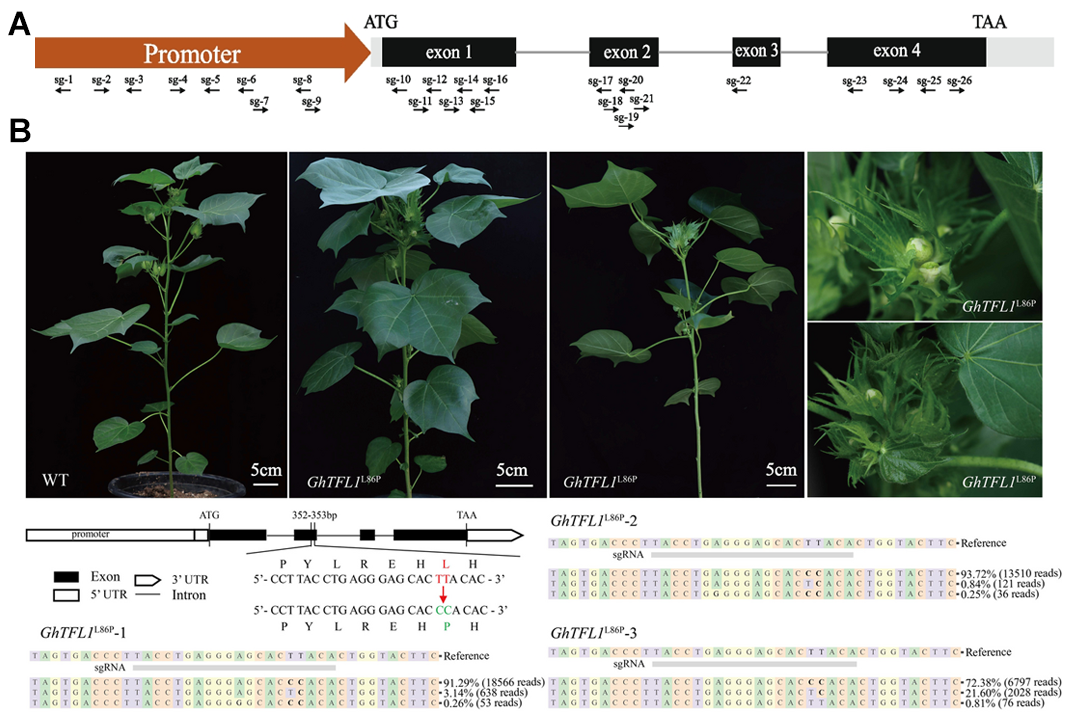
图4 GhTFL1L86P突变体表现出早开花、紧凑的花序表型(Wang et al., 2024)。(A)通过基因编辑靶向GhTFL1的大量位点;(B)GhTFL1L86P突变体表现出符合预期的理想表型。
上述三种方法都属于直接改造目的蛋白的结构,而现在所讲的第四种方法功能辅助调控法属于间接改造。其主要目的是将目的蛋白的辅因子进行改造,进而让目的蛋白表现出更强的功能。这里小远没有找到十分贴切的文献案例,以下的案例也能在一定程度上体现功能辅助调控法的可行性。
2020年10月,加州大学戴维斯分校的Jorge Dubcovsky课题组在Nature Biotechnology杂志上发表了一篇题为“A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants”的研究论文。作者发现,在辅助因子GIF 的存在下,GRF 对叶片增大的调控作用更加显著。基于这一观察,作者将小麦中的GRF和GIF1融合成一个蛋白。进一步研究显示,转化了GRF4-GIF1 的小麦,其再生效率显著提高,大幅缩短了小麦的转化时间(图5)。
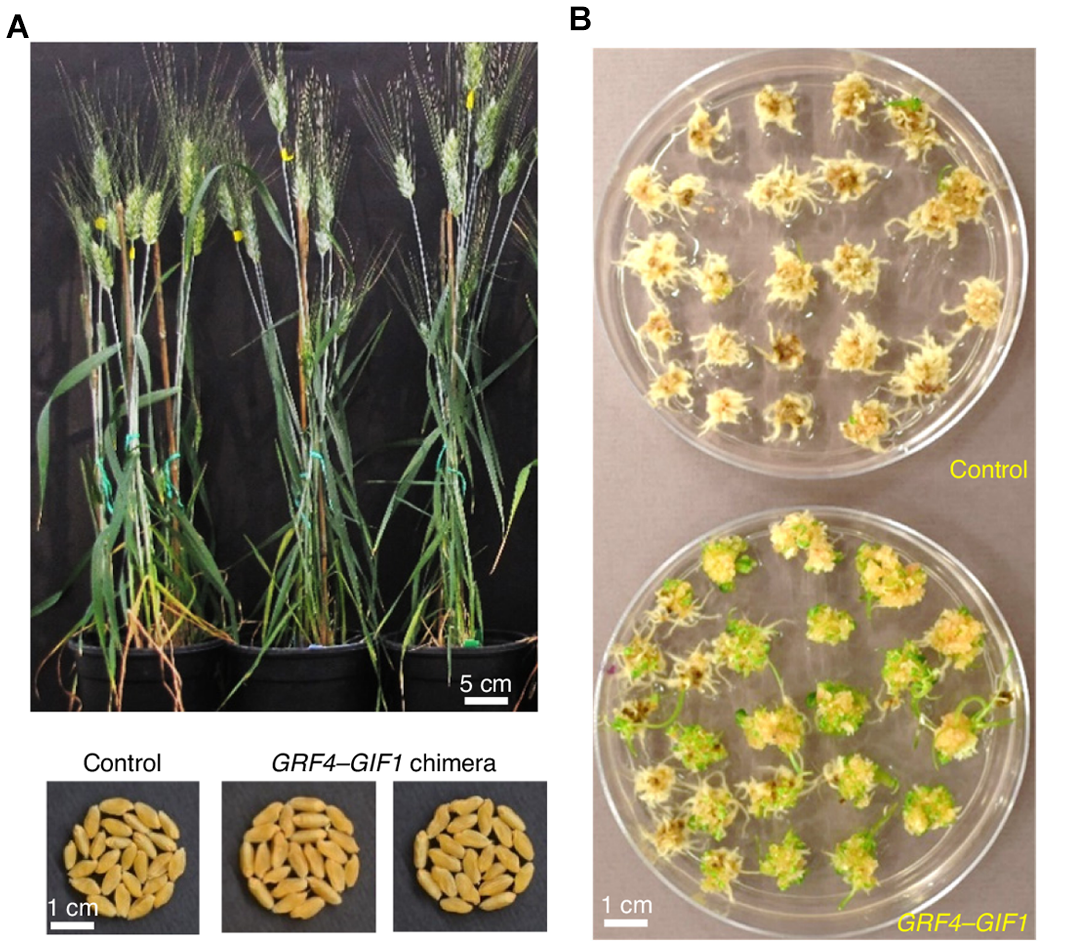
图5 转化了GRF4-GIF1 后能提高小麦的再生效率(Debernardi et al., 2020)。(A)转化了GRF4-GIF1 的小麦可育且表型正常;(B)转化了GRF4-GIF1 的小麦再生效率显著提高。
尽管该研究通过直接将辅助因子与目标蛋白融合实现了功能优化,但小远认为,通过改造辅助因子本身以增强其与目标蛋白的互作强度,或许也能达到类似的效果,为进一步提升转化效率提供了另一种可能性。
细心的读者可能已经注意到,前面提到的几种方法都离不开基因编辑工具的支持,这也从侧面印证了基因编辑技术的强大功能及其广泛的应用场景。在通过基因编辑获得突变体时,我们通常通过改变碱基序列,利用提前终止翻译或移码突变来实现目的蛋白的改造。虽然这些方法能够实现目标蛋白的突变,但需要注意的是,对于移码突变而言,虽然原有的目的蛋白失去了功能,但却可能生成一种新的蛋白质。而这种新蛋白质的功能往往被我们忽略。
那么,移码后产生的新蛋白质真的毫无作用吗?事实上并非如此。小远最近读到一篇有趣的文章,作者分享了他们在研究中意外发现,移码后产生的新蛋白质可能具备意想不到的功能。接下来,让我们一起来了解这一发现的细节。
2024年12月,浙江大学娄永根/李冉课题组在PNAS上发表了一篇题为“A frameshift mutation in JAZ10 resolves the growth versus defense dilemma in rice”的研究论文。在通过基因编辑研究水稻茉莉酸(JA)信号基因OsJAZ10 时,作者意外发现,提前终止翻译的突变体与野生型在表型上并无显著差异。然而,移码突变生成的突变体(命名为FJ10)却表现出增强的水稻生长能力和对褐飞虱的抗性特性。进一步研究还显示,FJ10 的过表达同样能产生类似的表型改良效果(图6)。通过深入分析,作者发现FJ10能与slender rice 1(OsSLR1)和F-box/Kelch 16(OsFBK16)发生相互作用。这些相互作用分别削弱了OsSLR1对赤霉素介导的生长抑制功能,以及OsFBK16对木质素介导防御反应的抑制作用。这种意外生成的FJ10蛋白,在优化水稻生长和防御之间建立了一个理想的平衡,展现了基因编辑生成新蛋白质潜在的育种应用价值。
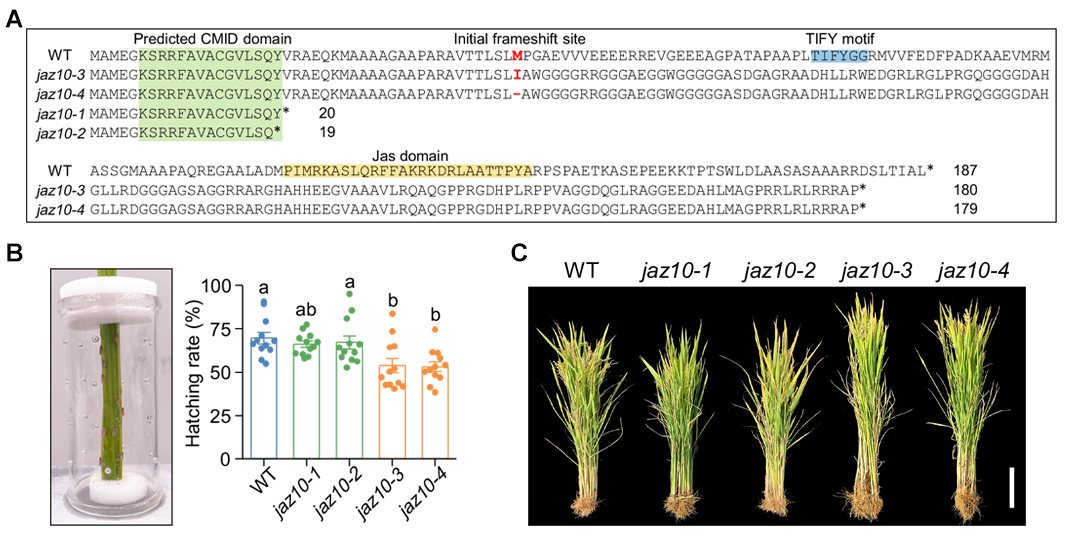
图6 基因编辑意外获得的移码材料FJ10表现出良好的生长和抗褐飞虱表型(Li et al., 2024)。(A)通过基因编辑获得了OsJAZ10 提前终止翻译的突变体和移码的突变体;(B)提前终止翻译的突变体jaz-1 和jaz-2 在生长和抗虫方面相比于野生型没有差异,移码突变体jaz-3 和jaz-4 在生长和抗虫方面表型优于野生型。
这个案例提醒我们,在通过基因编辑获得突变体时,需要特别关注因新蛋白产生而可能导致的不同表型。这种情况下,需要收集更多材料进行深入分析,以验证突变体表型的真实性和机制。当然,对于有心的研究者来说,不同表型的突变体也蕴藏着重要的研究价值,绝不会被轻易忽视。
植物蛋白结构改造尽管具有广阔的应用前景,但在实际研究和应用中仍面临诸多挑战和难点。首先,蛋白质的结构与功能关系极为复杂,而其结构又受到诸多外界因素的影响,例如辅助因子、化学环境等。这导致许多植物蛋白的结构尚未完全解析,使得预测关键结构域和功能性残基的难度增加,从而为定向改造带来了显著的障碍。其次,由于植物中许多蛋白的晶体结构数据仍然匮乏,尽管AlphaFold等工具能够提供预测支持,但在精度和可靠性方面仍存在一定的局限性,尤其是在多亚基复合物或特定环境下的预测中。此外,突变本身也可能引发意外的功能变化。例如,某些突变可能导致蛋白产生新的、未预料到的功能,这种“不可预测性”增加了实验设计的复杂性和风险。最后,尽管当前的基因编辑工具已经相当先进,但在一些特定场景下,仍难以实现对碱基序列的任意精准改动。这种技术局限性进一步限制了蛋白结构改造的范围和灵活性。
小远叨叨
在本文中,小远为大家介绍了蛋白结构改造的多种方法,希望能启发大家在进行蛋白功能研究时,更多地思考如何将研究成果应用于实际生产。有时候,仅靠基因的敲除或过表达无法满足实际需求,这就需要借助蛋白结构改造来实现特定的目标。尽管目前植物蛋白结构改造仍面临一些困难,但随着基因编辑技术的不断进步,以及植物蛋白晶体结构的逐步解析,这一过程将变得更加高效、精准和可控。期待未来蛋白结构改造能够在植物功能研究和实际应用中发挥更加重要的作用!
References:
Debernardi J M, Tricoli D M, Ercoli M F, et al. A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants[J]. Nature biotechnology, 2020, 38(11): 1274-1279.
Li L L, Xiao Y, Wang B, et al. A frameshift mutation in JAZ10 resolves the growth versus defense dilemma in rice[J]. Proceedings of the National Academy of Sciences, 2024, 121(52): e2413564121.
Li Z, Chen J, Liu C, et al. Natural variations of maize ZmLecRK1 determine its interaction with ZmBAK1 and resistance patterns to multiple pathogens[J]. Molecular Plant, 2024, 17(10): 1606-1623.
Wang G, Wang F, Xu Z, et al. Precise fine-turning of GhTFL1 by base editing tools defines ideal cotton plant architecture[J]. Genome Biology, 2024, 25(1): 59.
Zhu T, Wu X, Yuan G, et al. A resurfaced sensor NLR confers new recognition specificity to non-MAX effectors[J]. Journal of integrative plant biology, 2024.

