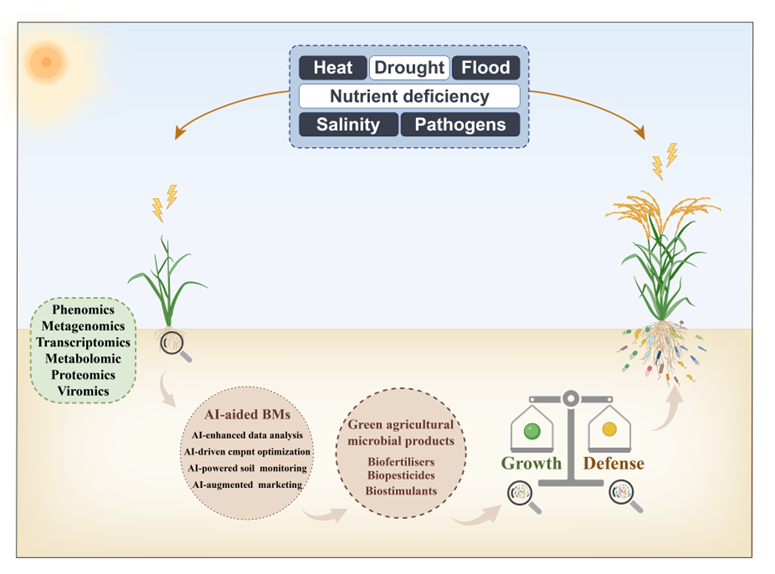
近日,Plant Biotechnology Journal 杂志在线发表了由姚燕来团队完成的综述论文——“Beneficial microorganisms: Regulating growth and defense for plant welfare”。文章从多个角度全面总结了植物益生微生物(Beneficial Microorganisms, BMs)在植物生长、抗病与抗逆等方面的重要作用,揭示了它们在植物生长-防御平衡中的调控机制,并探讨了其在农业可持续发展中的应用前景。

LRR-RLK对植物的生长、发育和防御等至关重要(图3a)。P. fluorescens能够特异性地诱导植物PSKR1(LRR-RLK家族)表达,促进植物生长,抑制免疫(Song et al., 2023)。X. oryzae产生PSY1类似物RaxX(Pruitt et al., 2017),PSY能够激活PSYR受体(LRR-RLK家族),低浓度PSY-PSYR激活拟南芥防御而非生长,高浓度PSY-PSYR激活拟南芥生长而非防御(Ogawa-Ohnishi et al., 2022)。
植物根际存在植物免疫抑制型(ISM)和免疫非抑制型(NSM)两类微生物群落(图3b)。当ISM的比例超过NSM时,根际微生物群落主要促进植物生长;反之,则增强植物免疫防御(Ma et al., 2021)。flg22是一种被广泛研究的MAMP,可以诱导植物防御相关基因表达。但BMs的flg22蛋白所激发的植物免疫反应较弱比病原菌弱(Trdá et al., 2014)。OsCERK1是一种参与MAMP级联的受体激酶,与AMF的短链几丁质结合后,会促进AMF与之共生;而与病原体的长链几丁质结合,则会激活植物免疫(Miyata et al., 2014;Wang et al., 2024)。OsCIE1能够调控OsCERK1的活性,以免植物出现过度免疫。当病原菌入侵时,OsCERK1磷酸化OsCIE1阻断其活性,产生强烈的免疫反应。病原菌被清除后,OsCIE1则会重新泛素化OsCERK1降低其活性,并恢复免疫平衡。上述均展现了BMs可以保护植物因过度免疫而损害自身生长。
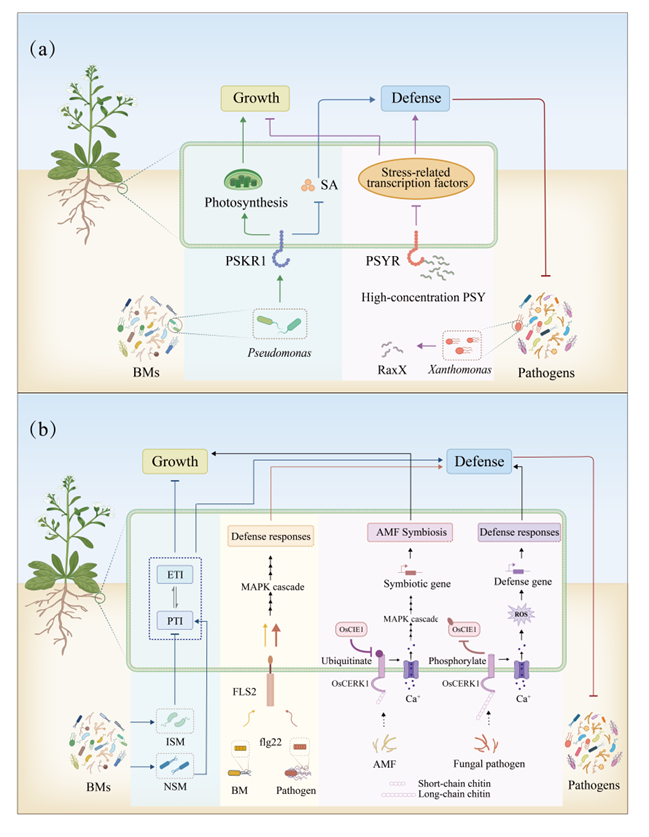
图3 BMs调控生物胁迫下植物的生长-防御权衡
干旱下,ABA参与生长调节和防御激活体现了经典的生长-防御权衡,即资源分配给一个功能则可能会削弱另一个功能(图4a)。多种BMs被报道能够通过ABA途径调控植物的抗旱能力,如AMF、P. indica、A. brasilense、 S. pactum、P. argentinensis SA190、B. cereus NJ01等。然而,其内部识别相应的分子机制还需进一步探究。
MYC2是植物中的一种关键转录因子,能够调控植物生长-防御权衡。在正常光照下,BMs调控MYC2来增强植物的生长和免疫防御;而在低光照下,BMs则主要促进生长而非防御(Hou et al., 2021)(图4b)。然而BMs调控MYC2的分子机制仍然未知。PHR1是植物体内磷信号转导的主要转录因子,在基于磷可用性的生长-防御权衡中至关重要。磷缺乏下,B. amyloliquefaciens GB03产生二乙酰(DA)(Morcillo et al., 2020),增强植物免疫反应,进而增强植物对磷酸盐缺乏的超敏感性;磷充足下,DA则抑制植物的ROS产生,而不影响植物抗病性(图4c)。这些研究对于优化不同磷条件下的植物适应性至关重要。
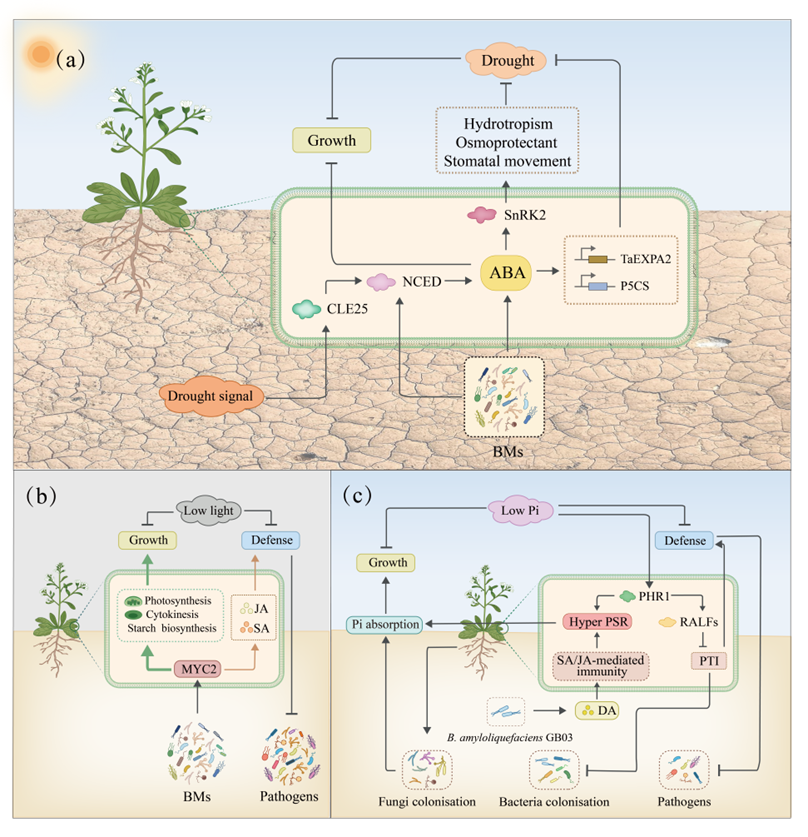
图4 BMs调控非生物胁迫下植物的生长-防御权衡
He, Z., Webster, S., and He, S. Y. (2022). Growth-defense trade-offs in plants. Curr. Biol. 32, R634–R639.
Hou, S., Thiergart, T., Vannier, N., Mesny, F., Ziegler, J., Pickel, B., and Hacquard, S. (2021). A microbiota-root-shoot circuit favours Arabidopsis growth over defence under suboptimal light. Nat. Plants. 7, 1078–1092.
Ma, K. W., Niu, Y., Jia, Y., Ordon, J., Copeland, C., Emonet, A., Geldner, N., Guan, R., Stolze, S. C., Nakagami, H., et al. (2021). Coordination of microbe-host homeostasis by crosstalk with plant innate immunity. Nat. Plants. 7, 814–825.
Morcillo, R. J., Singh, S. K., He, D., An, G., Vílchez, J. I., Tang, K., Yuan, F., Sun, Y., Shao, C., Zhang, S., et al. (2020). Rhizobacterium‐derived diacetyl modulates plant immunity in a phosphate‐dependent manner. EMBO J. 39, e102602.
Ogawa-Ohnishi, M., Yamashita, T., Kakita, M., Nakayama, T., Ohkubo, Y., Hayashi, Y., Yamashita, Y., Nomura, T., Noda, S., Shinohara, H., et al. (2022). Peptide ligand-mediated trade-off between plant growth and stress response. Science. 378, 175–180.
Pruitt, R. N., Joe, A., Zhang, W., Feng, W., Stewart, V., Schwessinger, B., Dinneny, J. R., and Ronald, P. C. (2017). A microbially derived tyrosine-sulfated peptide mimics a plant peptide hormone. New Phytol. 215, 725–736.
Song, S., Morales Moreira, Z., Briggs, A. L., Zhang, X.-C., Diener, A. C., and Haney, C. H. (2023). PSKR1 balances the plant growth-defence trade-off in the rhizosphere microbiome. Nat. Plants. 9, 2071–2084.
Trdá, L., Fernandez, O., Boutrot, F., Héloir, M. C., Kelloniemi, J., Daire, X., Adrian, M., Clément, C., Zipfel, C., Dorey, S., et al. (2014). The grapevine flagellin receptor VvFLS2 differentially recognizes flagellin-derived epitopes from the endophytic growth-promoting bacterium Burkholderia phytofirmans and plant pathogenic bacteria. New Phytol. 201, 1371–1384.
Wang, G., Chen, X., Yu, C., Shi, X., Lan, W., Gao, C., Yang, J., Dai, H., Zhang, X., Zhang, H., et al. (2024). Release of a ubiquitin brake activates OsCERK1-triggered immunity in rice. Nature. 629, 1158–1164.
图文来源:植物生物技术Pbj
