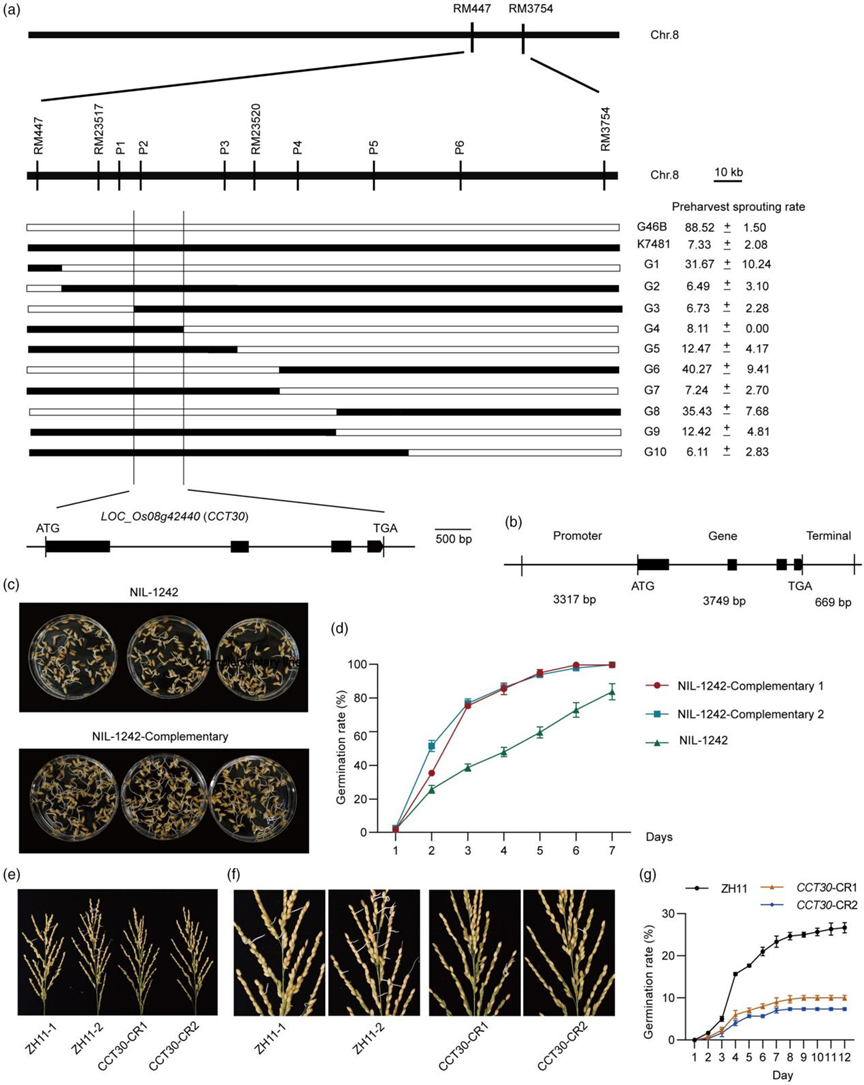
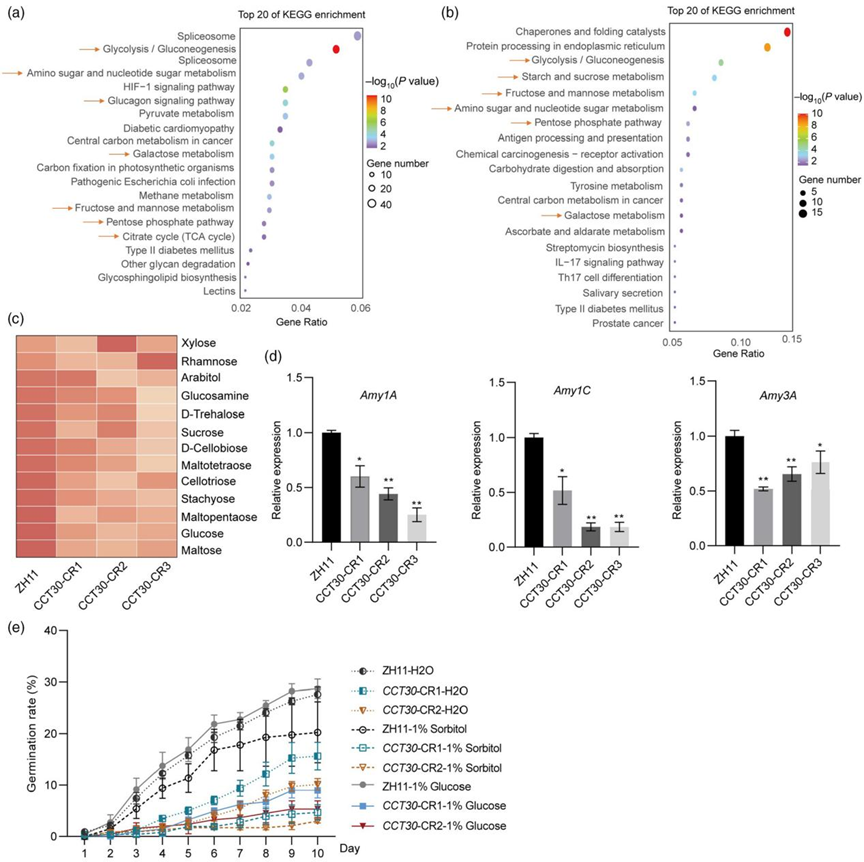
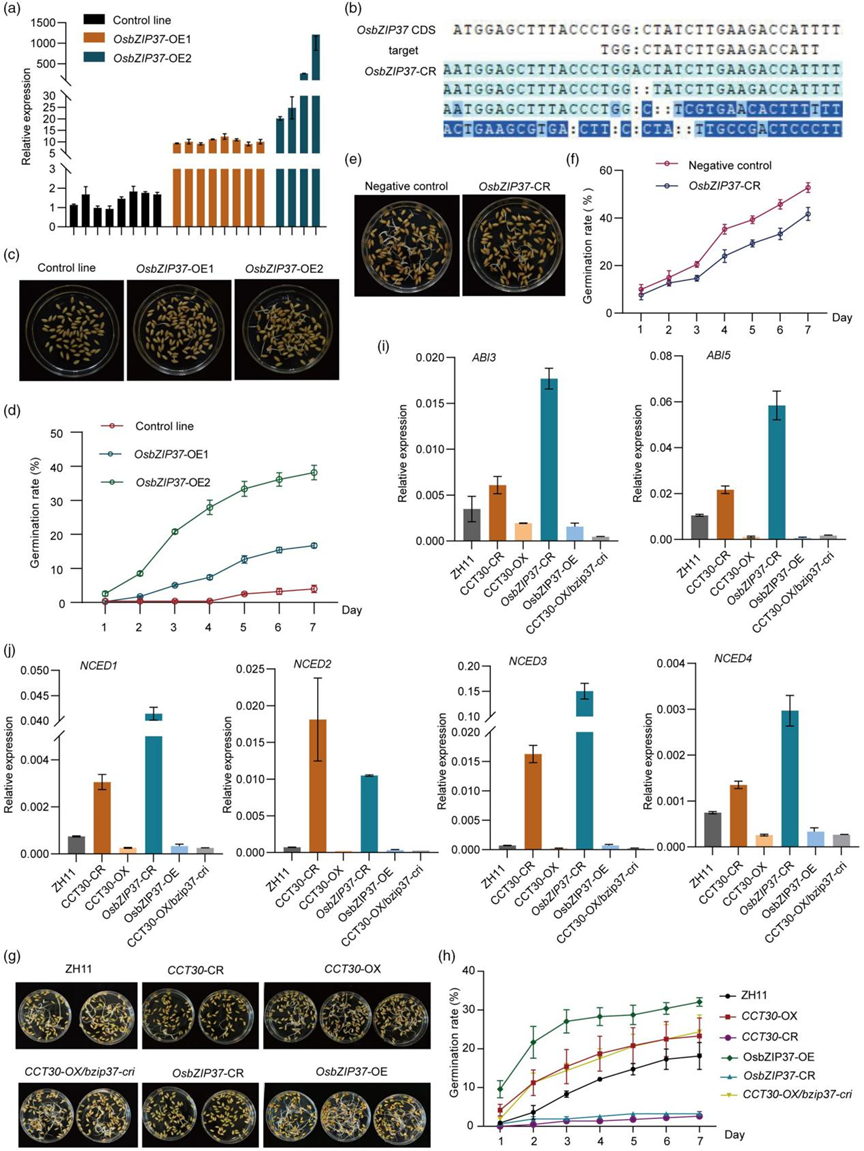
近日,Plant Biotechnology Journal 杂志在线发表了由华中农业大学作物遗传改良全国重点实验室、崖州湾国家实验室邢永忠教授课题组联合四川省农业科学院作物研究所任光俊研究员团队撰写的“The transcription factor CCT30 promotes rice preharvest sprouting by regulating sugar signalling to inhibit the ABA-mediated pathway”论文。该研究通过对前期鉴定到的调控水稻穗发芽的主效QTL qPSR8 进行精细定位(Gao et al., 2008),克隆了该QTL位点的候选基因CCT30。研究者通过对CCT30 转基因材料休眠表型的鉴定,证实了CCT30 促进水稻穗发芽的新功能。进一步的ABA含量及可溶性糖含量测定、外源ABA及葡萄糖处理分析,以及ABA通路和糖代谢相关基因表达量检测等,明确了CCT30通过增强糖信号、抑制ABA的合成及信号途径从而减弱种子的休眠、促进水稻穗发芽。此外,研究发现CCT30与转录因子OsbZIP37相互作用,它们协同调控下游基因,负向调节水稻种子休眠。研究团队报道的这一关于水稻穗发芽调控基因CCT30克隆及机制解析的工作,为培育水稻抗穗发芽品种提供了重要基因和理论基础。


l An, L., Tao, Y., Chen, H., He, M., Xiao, F., Li, G., Ding, Y. et al. (2020) Embryo-endosperm interaction and its agronomic relevance to rice quality. Front.Plant Sci. 11, 587641.
l Farooq, M.A., Ma, W., Shen, S. and Gu, A. (2022) Underlying biochemical andmolecular mechanisms for seed germination. Int. J. Mol. Sci. 23, 8502.
l Finch- Savage, W.E. and Leubner-Metzger, G. (2006) Seed dormancy and thecontrol of germination. New Phytol. 171, 501–523.
l Gao, F., Ren, G., Lu, X., Sun, S., Li, G., Gao, Y., Luo.t al. (2008) QTL analysisfor resistance to preharvest sprouting in rice (Oryza sativa). Plant Breed. 127,268–273
l Gubler, F., Millar, A.A. and Jacobsen, J.V. (2005) Dormancy release, ABA andpre- harvest sprouting. Curr. Opin. Plant Biol. 8, 183–187.
l Pan, T., Lin, L., Wang, J., Liu, Q. and Wei, C. (2018) Long branch- chains ofamylopectin with B-type crystallinity in rice seed with inhibition of starchbranching enzyme I and IIb resist in situ degradation and inhibit plant growthduring seedling development. BMC Plant Biol. 18, 1–11.
l Shu, K., Liu, X.D., Xie, Q. and He, Z.H. (2016) Two faces of one seed:hormonal regulation of dormancy and germination. Mol. Plant, 9, 34–45.
l Sohn, S.I., Pandian, S., Kumar, T.S., Zoclanclounon, Y.A.B., Muthuramalingam,P., Shilpha, J., Satish, L. et al. (2021) Seed dormancy and pre- harvestsprouting in rice-an updated overview. Int. J. Mol. Sci. 22, 11804.
l Tai, L., Wang, H.J., Xu, X.J., Sun, W.H., Ju, L., Liu, W.T., Li, W.Q. et al. (2021)Pre- harvest sprouting in cereals: genetic and biochemical mechanisms. J. Exp.Bot. 72, 2857–2876.
l Tao, Y., An, L., Xiao, F., Li, G., Ding, Y., Paul, M.J. and Liu, Z. (2022) Integrationof embryo–endosperm interaction into a holistic and dynamic picture of seeddevelopment using a rice mutant with notched-belly kernels. Crop J. 10, 729–742.
图文来源:植物生物技术Pbj
