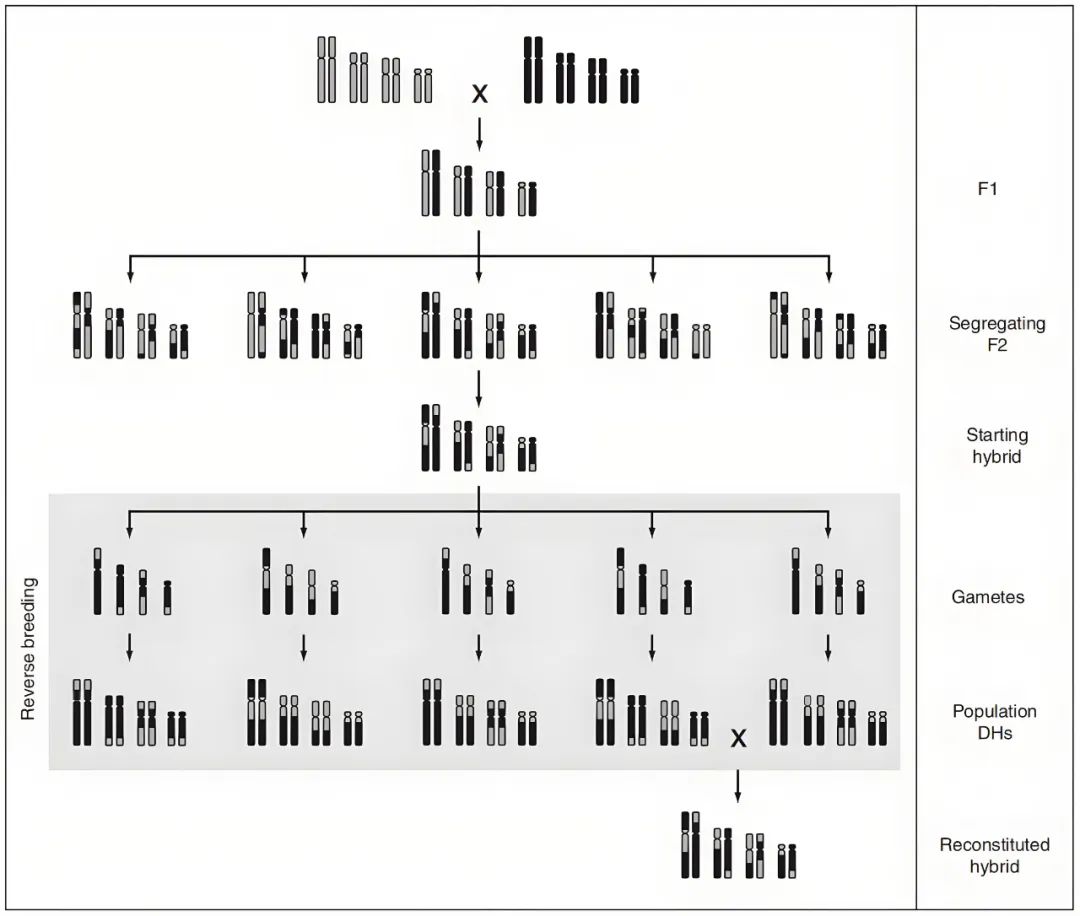
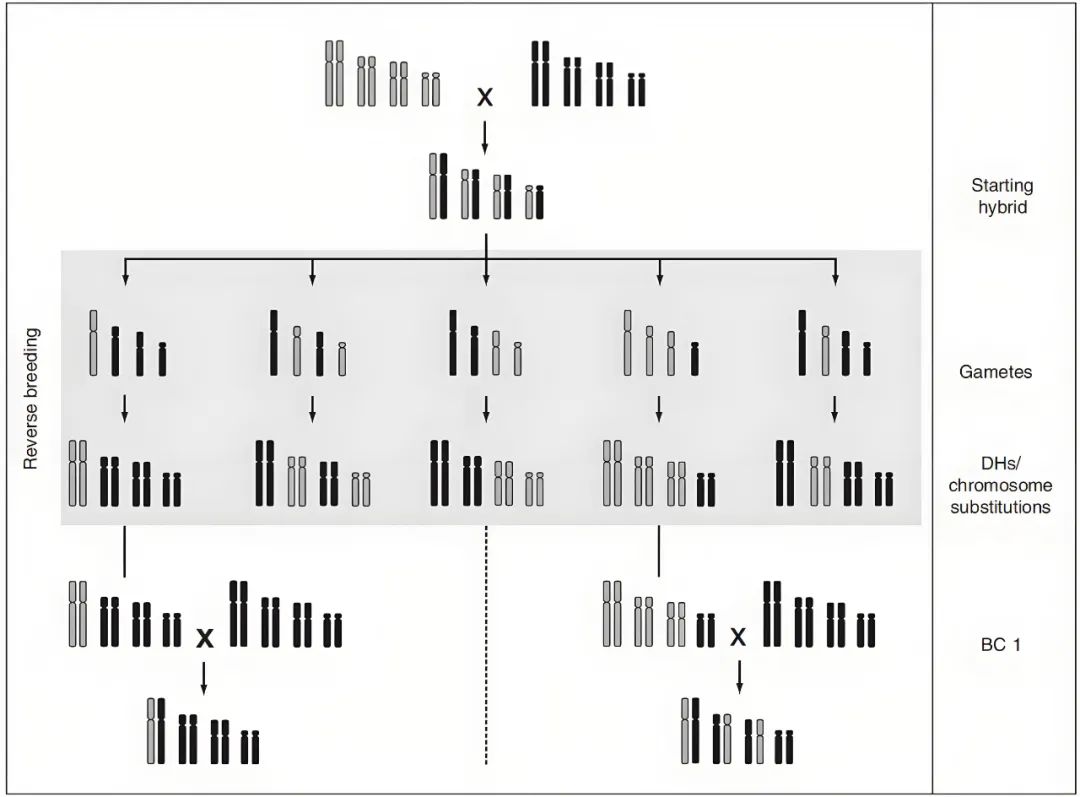
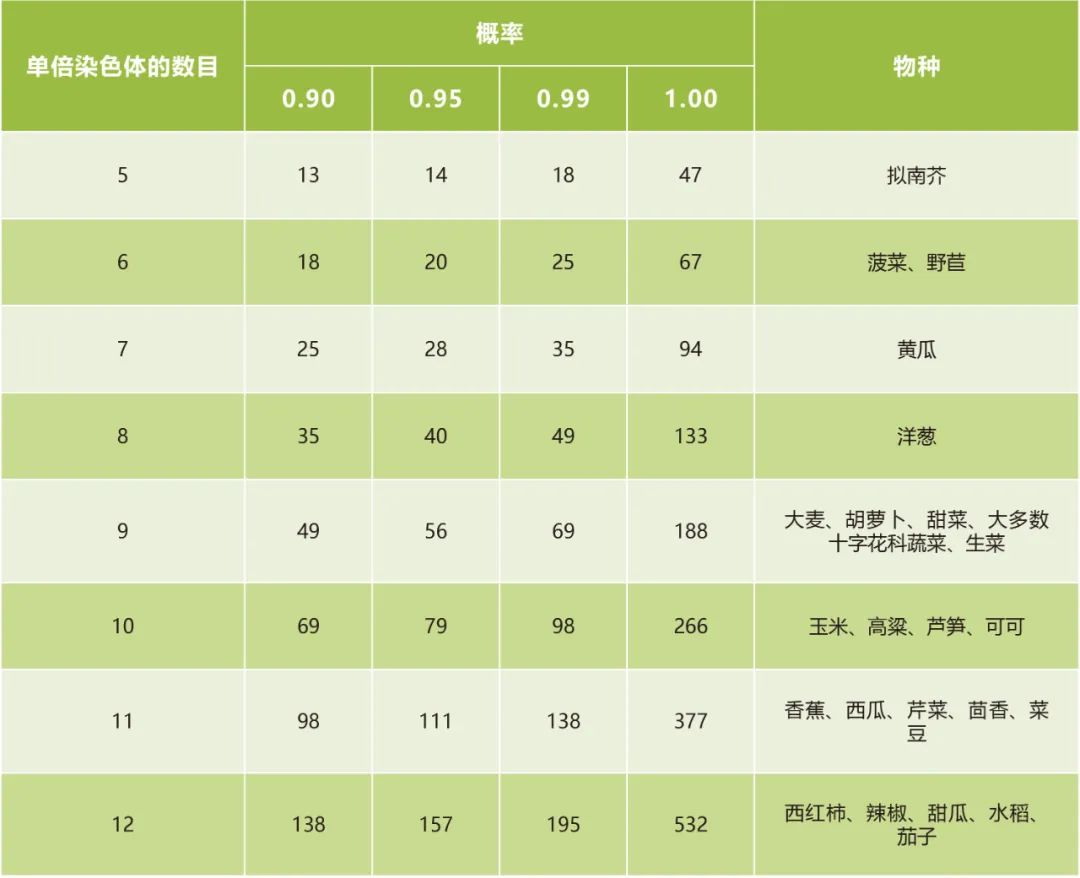
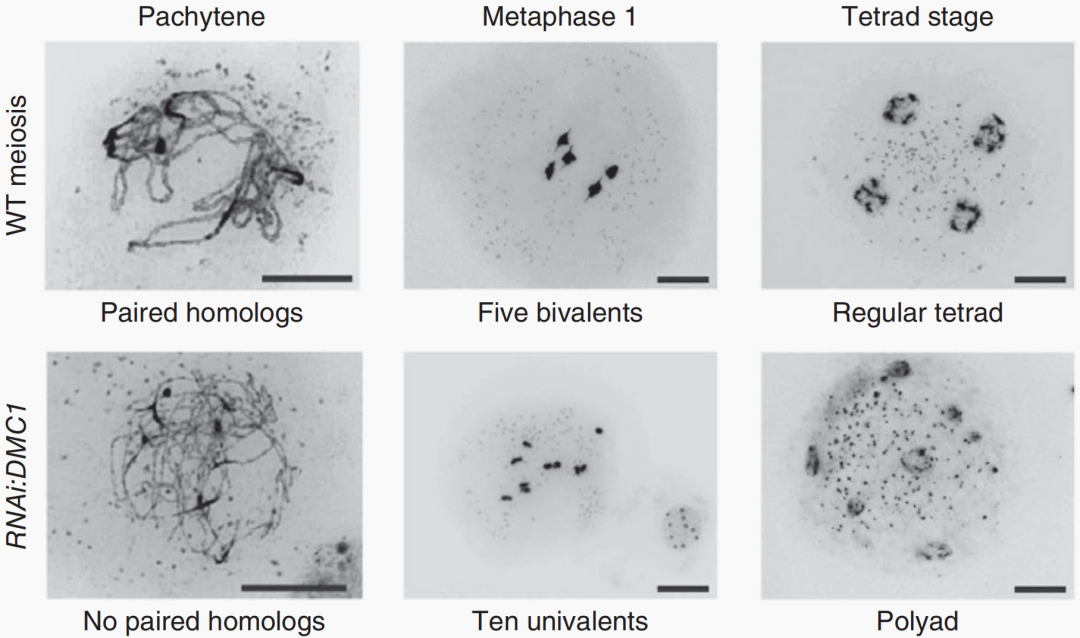
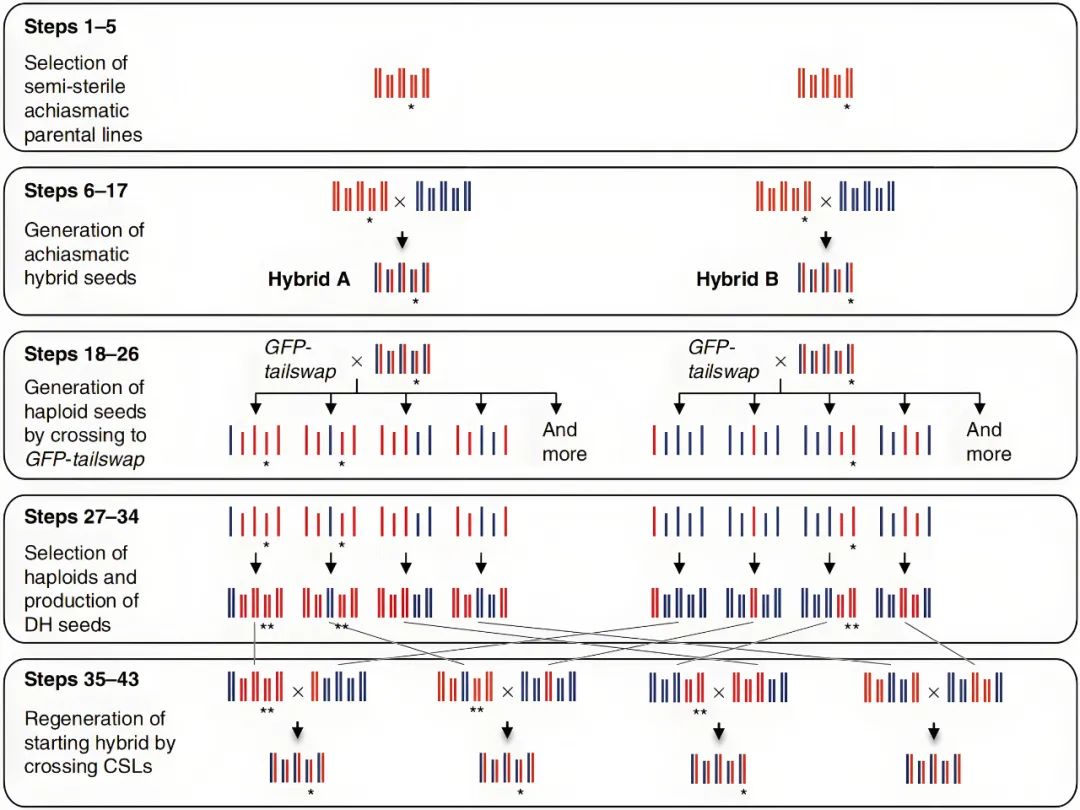
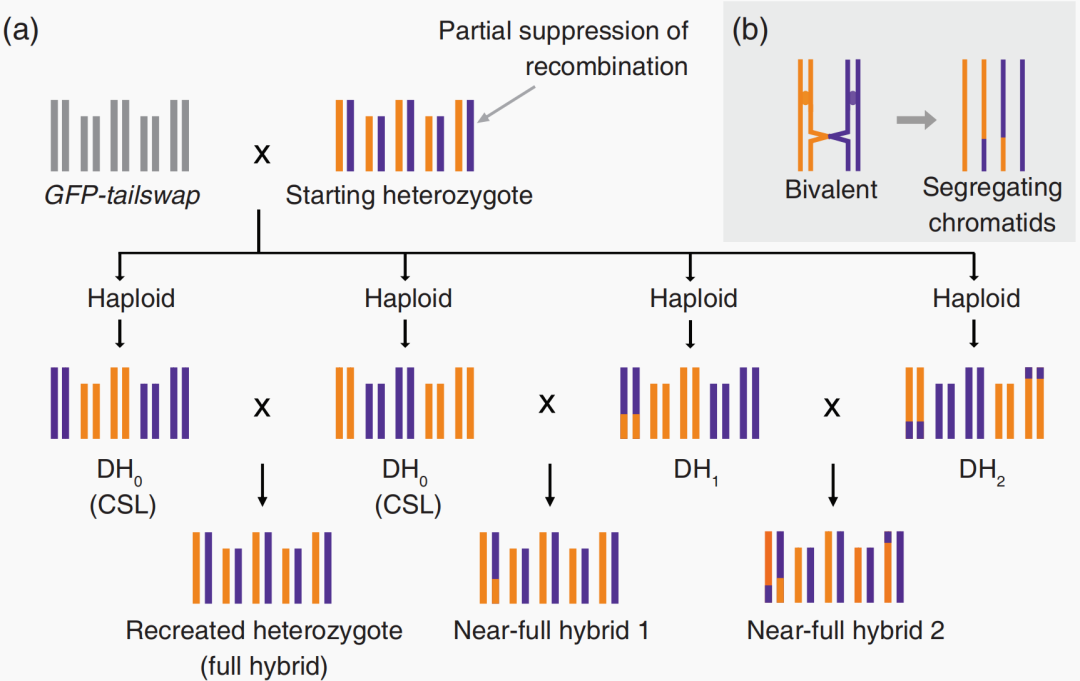
图11 通过抑制重组和产生DHs来对拟南芥进行反向育种(Calvo-Baltanas et al., 2020)。选择一个杂合子作为起始材料,这个杂合子的交换受到抑制,用其花粉为GFP-tailswap植株授粉,产生单倍体及DHs用于重建起始杂交种。
反向育种技术需要结合分子标记技术以辅助筛选。2015年,南通大学李平团队在Journal of Integrative Agriculture杂志上发表了一篇题为“Development and application of marker-assisted reverse breeding using hybrid maize germplasm”的研究论文,作者利用分子标记从一个亲本信息未知的玉米起始杂交种筛选出了与相应父本和母本高度相似的材料。由于玉米种皮基因全部来自母本,而胚的基因一半来自母本、一半来自父本,分别获取起始杂交种种皮和胚的DNA,利用高密度芯片推导出双亲的基因型,再选择均匀分布于10条染色体的192个双亲间的多态性标记构建低密度芯片用于后续的辅助筛选。作者从F2代杂交种开始,经过三轮的标记辅助筛选F3、F4和F5代,获得了与亲本基因型有80%相同的品系(图12)。该研究使用的并非是前述Erik Wijnker提出的反向育种思路,因此小远在这里并未将其列入“实现玉米反向育种技术”之列,这里是提示大家可以在反向育种的实施路径中采用此方法以提高筛选效率。

图12 通过分子标记辅助反向育种开发的近交品系,其基因型和表型与杂交种的父母本相似(Yi-Xin G et al., 2015)。
小远叨叨
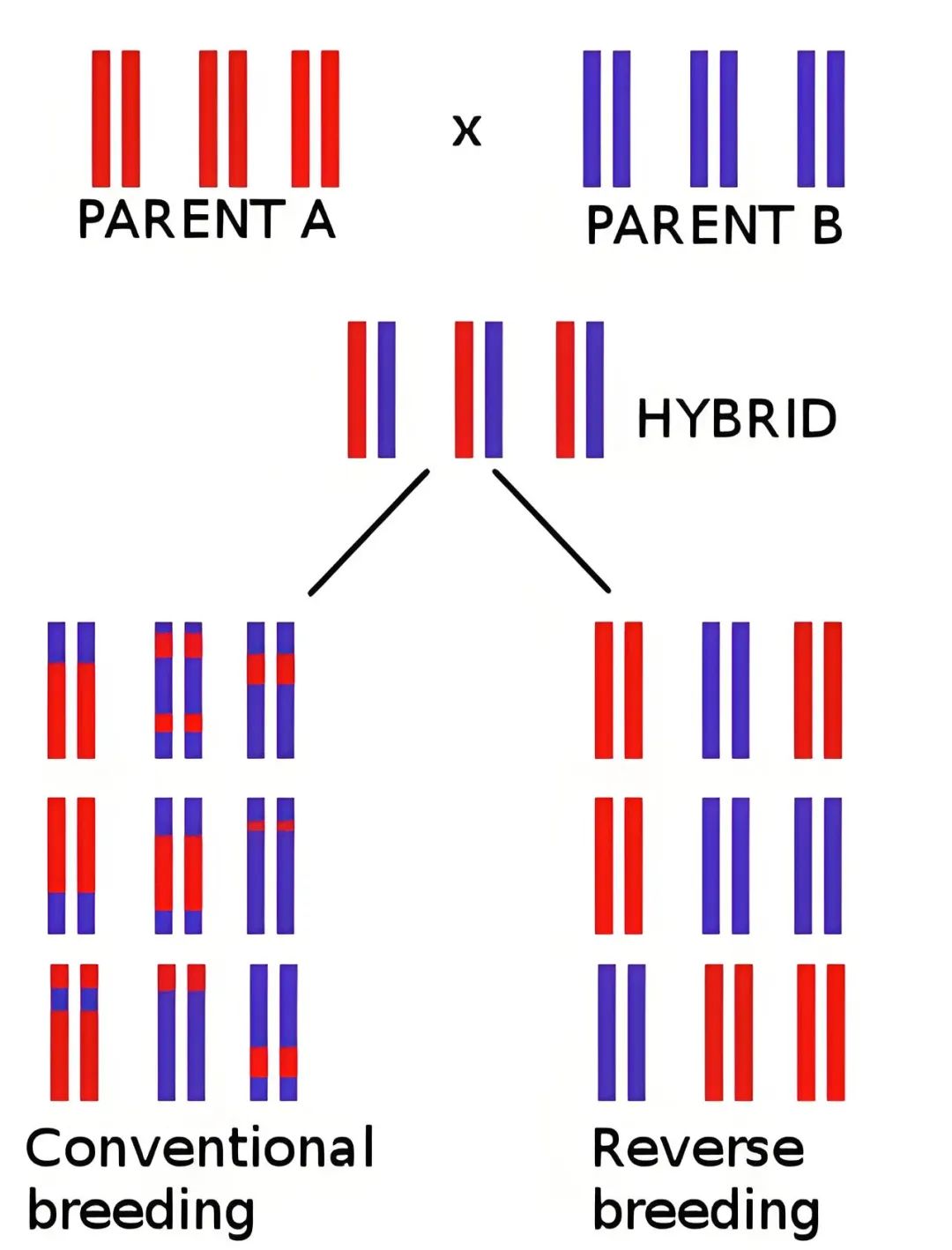
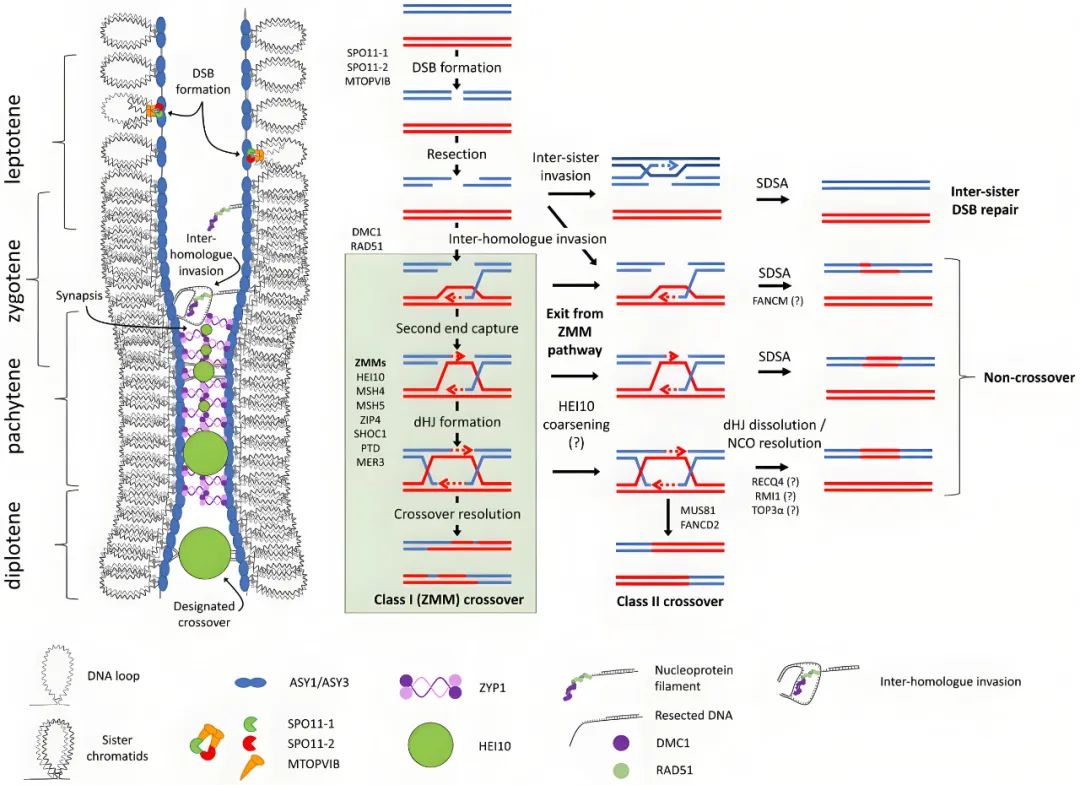
看完关于反向育种的相关资料,伯小远觉得这个技术如果做成功了,对做杂交育种的公司可能会有很大的冲击。伯小远发现反向育种似乎在十几年前研究的比较多,2012年反向育种在拟南芥中做出来之后,后面那么多年为什么没有在作物中做出来呢?伯小远觉得可能有三个原因,一个是关于作物中减数分裂的基础研究可能做的还不够成熟,二是当时的单倍体诱导技术还不够成熟,另外该方法也受物种的遗传转化限制。但现在再看这三点,基础研究发展的很迅速,减数分裂基因、单倍体诱导基因被发现的越来越多,基因编辑技术、单倍体诱导技术都在大力发展,甚至连破除遗传转化限制的HI-Edit、IMGE以及嫁接技术也有了,那么,在不久的将来反向育种会不会在作物中也实现呢?大家可以来一起讨论喔!
Calvo-Baltanas V, Wijnen CL, Yang C, Lukhovitskaya N, de Snoo CB, Hohenwarter L, Keurentjes J, de Jong H, Schnittger A, Wijnker E (2020) Meiotic crossover reduction by virus-induced gene silencing enables the efficient generation of chromosome substitution lines and reverse breeding in Arabidopsis thaliana. PLANT J 104: 1437-1452
Dirks R, van Dun K, de Snoo CB, van den Berg M, Lelivelt CL, Voermans W, Woudenberg L, de Wit JP, Reinink K, Schut JW, van der Zeeuw E, Vogelaar A, Freymark G, Gutteling EW, Keppel MN, van Drongelen P, Kieny M, Ellul P, Touraev A, Ma H, de Jong H, Wijnker E (2009) Reverse breeding: a novel breeding approach based on engineered meiosis. PLANT BIOTECHNOL J 7: 837-845
Dupré, A., Boyer-Chatenet, L., Sattler, R. M., Modi, A. P., Lee, J. H., Nicolette, M. L., … &Gautier, J. (2008). A forward chemical genetic screen reveals an inhibitor of the Mre11-Rad50-Nbs1 complex. Nature chemical biology, 4(2), 119-125.
Huang Y, Meng X, Rao Y, Xie Y, Sun T, Chen W, Wei X, Xiong J, Yu H, Li J, Wang K (2024) OsWUS-driven synthetic apomixis in hybrid rice. PLANT COMMUN: 101136
Khanday I, Skinner D, Yang B, Mercier R, Sundaresan V (2019) A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. NATURE 565: 91-95
Marimuthu MP, Jolivet S, Ravi M, Pereira L, Davda JN, Cromer L, Wang L, Nogue F, Chan SW, Siddiqi I, Mercier R (2011) Synthetic clonal reproduction through seeds. SCIENCE 331: 876
Song M, Wang W, Ji C, Li S, Liu W, Hu X, Feng A, Ruan S, Du S, Wang H, Dai K, Guo L, Qian Q, Si H, Hu X (2024) Simultaneous production of high-frequency synthetic apomixis with high fertility and improved agronomic traits in hybrid rice. MOL PLANT 17: 4-7
Wang C, Liu Q, Shen Y, Hua Y, Wang J, Lin J, Wu M, Sun T, Cheng Z, Mercier R, Wang K (2019) Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes. NAT BIOTECHNOL 37: 283-286
Wijnker E, Deurhof L, van de Belt J, de Snoo CB, Blankestijn H, Becker F, Ravi M, Chan SW, van Dun K, Lelivelt CL, de Jong H, Dirks R, Keurentjes JJ (2014) Hybrid recreation by reverse breeding in Arabidopsis thaliana. NAT PROTOC 9: 761-772
Yi-Xin, G. U. A. N., Wang, B. H., Yan, F. E. N. G., & Ping, L. I. (2015). Development and application of marker-assisted reverse breeding using hybrid maize germplasm. Journal of Integrative Agriculture, 14(12), 2538-2546.
Wijnker E, van Dun K, de Snoo CB, Lelivelt CL, Keurentjes JJ, Naharudin NS, Ravi M, Chan SW, de Jong H, Dirks R (2012) Reverse breeding in Arabidopsis thaliana generates homozygous parental lines from a heterozygous plant. NAT GENET 44: 467-470