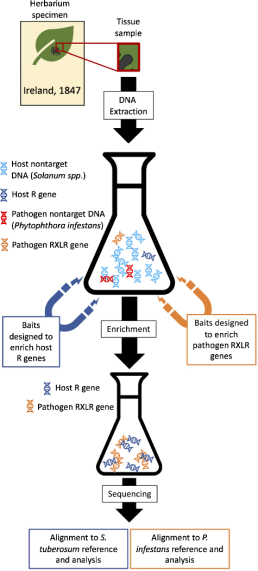
疫霉是一种主要的卵菌植物病原体,引起马铃薯晚疫病,导致1845-1852年爱尔兰马铃薯饥荒.近日,来自美国北卡州立大学昆虫学和植物病理学学系的Jean Beagle Ristaino团队在Nature Communications发表研究成果“Evolution of Phytophthora infestans on its potato host since the Irish potato famine”,该研究揭示了在自然选择和人工选择的作用下,饥荒以来病原菌毒力基因和宿主抗性基因发生了显著变化.图1 样品处理的图像大纲.首先,作者利用靶向富集测序,同时对1845-1954年植物标本室感染茄属植物的宿主R基因(可识别RXLR效应蛋白)和病原体效应子进行测序,同时分析和记录病原体对抗性基因部署的变化,此外调查自爱尔兰饥荒以来R基因和RXLR基因丰度的时间变化,最后评估采样期间R基因和RXLR效应基因的进化和多样性(图1).图2 Avr1和R1的氨基酸序列历史上的FAM-1谱系(导致饥荒的感染疟原虫的克隆系,在20世纪30-50年代被US-1系取代)具有毒性的Avr1等位基因,并在育种者应用于马铃薯之前破坏R1抗性基因的能力.测序结果表明,所有样品中,无论宿主R1基因的变异如何,都显示出相同的Avr1序列(图2),这表明R1对效应体Avr1的选择不足.FAM-1和US-1谱系的所有样品都具有可被R1识别的AL毒性抗性断裂等位基因(图2A).图3 疫霉菌1306参考基因组中所有诱饵RXLR位点的覆盖率.
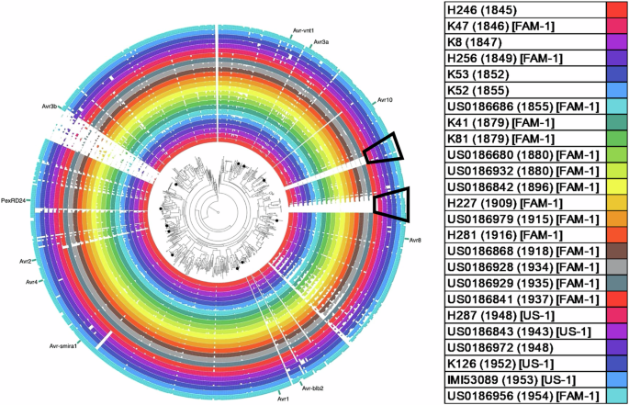
历史致病疫霉的RXLR基因组显示出总体稳定性,直到US-1谱系出现(在1948年的样品组中),此时观察到许多附加效应子的基因组扩增(图3)图4马铃薯SolTub3.0参考基因组中所有诱饵R位点的覆盖率
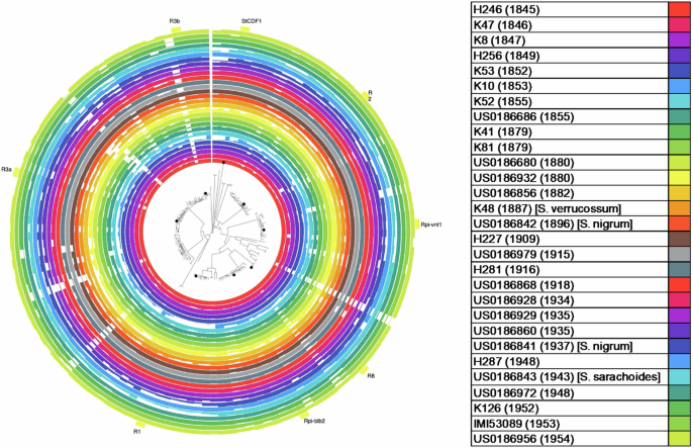
对于FAM-1谱系,平均发现664个覆盖效应子,相比之下,US-1谱系平均发现675个覆盖效应子.茄属宿主R基因组中的覆盖范围随着时间的推移似乎更易变化,不同基因座之间的趋势不太明显,这反映了马铃薯随着时间推移的不同育种努力(图4).图5 按照样品收集的时间45分类的所有RXLR基因的选择统计的小提琴图.
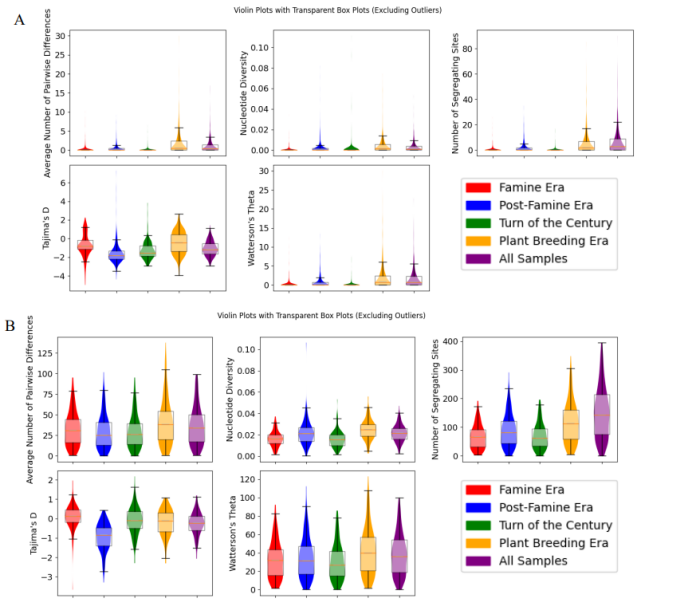
同时对每个采样期间的样本群组进行核苷酸多样性统计:饥荒期(1845-1852)、饥荒后期(1853-1883)、世纪之交(1884-1924)、植物育种期(1925-1954)以及总体(图5).RXLR基因在植物育种期的成对差异数量、核苷酸多样性和分离位点数量均高于早期.Tajima的D在整个采样期间(所有四个时代)的RXLR基因组始终为阴性,表明数据集中存在大量罕见的罕见多态性.这可能是由于出现瓶颈期后种群规模的扩张,或对RXLR基因组的正向选择.R基因组的Tajima D在饥荒后时期最低,表明在该时期存在负选择压力(图5B).综上:作者利用植物标本上的效应基因和R基因的定向测序,研究了1845-1954年间马铃薯与病原菌的联合进化.目前相关的效应物历史上存在于病原菌中,但与现代参考基因组相比,具有替代等位基因.历史上的FAM-1谱系具有有毒的Avr1等位基因,并且能够在育种者将R1抗性基因应用于马铃薯之前破坏R1抗性基因.FAM-1谱系是二倍体,但后来出现了三倍体US-1谱系.在自然选择和人工选择的作用下,饥荒以来病原菌毒力基因和宿主抗性基因发生了显著变化.
原文链接: