本文内容速览:

研究植物基因的上下游调控关系,对于理解植物生长发育、抗逆性和遗传改良等方面的机制具有重要意义。在“辨清敌我,统一战线——确定基因上下游关系”一文中,小远已经为大家讲解了确定基因上下游关系的方法,包括证明某信号分子参与调控某信号通路、在转基因材料中确定基因之间的调控关系,以及在没有转基因材料的情况下如何确定基因之间的调控关系。然而,很多小伙伴对利用转基因材料来探究基因的上下游关系还存在一些疑问,为了帮助大家更好地理解这个问题,小远再次搜集了一些文献进行详细的讲解,希望这些信息能为大家提供更多的帮助!
假设基因A具有正调控作用,基因B具有负调控作用。首先分别创制基因A和基因B的单突变体,比较A突变体与野生型的表型差异,确定基因A的正调控作用;比较B突变体与野生型的表型差异,确定基因B的负调控作用。然后创制双突变体,比较双突变体A/B与单突变体A和B的表型差异,分析基因A和B之间的调控关系:
如果A突变体的表型是某些特定性状缺失或减弱,而双突变体A/B的表型与A突变体相似,说明基因B的突变不能恢复基因A突变导致的性状缺失或减弱,这种情况表明基因B在基因A的上游。如案例1所示。
如果A突变体的表型是某些特定性状缺失或减弱,而双突变体A/B的表型与B突变体相似,说明基因B的突变能够恢复基因A突变导致的性状缺失或减弱,这种情况表明基因A在基因B的上游。如案例2所示。
案例1
2024年7月,中国农业大学张小兰课题组在The Plant Cell杂志上发表了一篇题为“The AGAMOUS-LIKE 16-GENERAL REGULATORY FACTOR 1 module regulates axillary bud outgrowth via catabolism of abscisic acid in cucumber”的研究论文。作者发现黄瓜MADS-box转录因子AGAMOUS-LIKE 16(CsAGL16)正调控黄瓜侧枝伸长,CsAGL16突变后导致侧枝减少,而过表达则促进了侧枝伸长。此外,作者还发现黄瓜General Regulatory Factor 1(CsGRF1)与CsAGL16在蛋白水平上相互作用,并抑制了CsAGL16对CsCYP707A4的转录激活。CsGRF1负调控黄瓜侧枝伸长,CsGRF1突变后促进了腋芽伸长并降低了腋芽中的ABA水平。为了探究CsGRF1和CsAGL16的遗传关系,作者创制了Csagl16 Csgrf1双突变体,发现Csagl16 Csgrf1双突变体表现出与Csagl16单突变体相似的分枝表型(图1A、B),与WI1983M和Csgrf1相比,Csagl16单突变体和Csagl16 Csgrf1双突变体每株的平均侧枝数量和分枝总长度均显著减少(图1C、D)。这些结果表明在黄瓜侧枝发育过程中,CsGRF1在CsAGL16上游起作用。

图1 Csagl16 Csgrf1双突变体的表型分析(Chen et al., 2024)。(A)WI1983M、Csagl16、Csgrf1、Csagl16 Csgrf1植株的表型;(B)WI1983M、Csagl16、Csgrf1、Csagl16 Csgrf1植株的侧枝位置和长度。每列表示一株黄瓜植株。每层代表黄瓜植株中的一个节点。绿色方块:无芽生长;黄色方块:芽长≥4cm;棕色方块:侧枝长≥50cm;(C)WI1983M、Csagl16、Csgrf1、Csagl16 Csgrf1植株的单株平均分枝数;(D)WI1983M、Csagl16、Csgrf1、Csagl16 Csgrf1植株的单株侧枝总长度;(E)CsCYP70A4在WI1983M、Csagl16、Csgrf1、Csagl16 Csgrf1植株腋芽中的相对表达量。
案例2
2022年6月,中科院分子植物科学卓越创新中心林鸿宣课题组与上海交通大学林尤舜课题组联合在Science杂志上发表了一篇题为“A genetic module at one locus in rice protects chloroplasts to enhance thermotolerance”的研究论文。作者定位克隆到一个控制水稻高温抗性的新QTL位点TT3。非洲栽培稻(CG14)来源的TT3相较于亚洲栽培稻(WYJ)来源的TT3具有更强的高温抗性。进一步研究发现,TT3位点中存在两个拮抗调控水稻高温抗性的QTL基因TT3.1和TT3.2,其中TT3.1正向调控高温抗性,而TT3.2负向调控高温抗性(图2)。将tt3.1突变体、tt3.2突变体杂交后得到tt3.1 tt3.2双突变体,检测tt3.1 tt3.2双突变体在正常和42℃高温条件下生长7天后的表型和存活率,发现tt3.1 tt3.2双突变体的表型与tt3.2突变体表型相似(图3),说明TT3.1在TT3.2的上游起作用。

图2 TT3.1与TT3.2过表达或敲除株系的耐热表型分析(Zhang et al., 2022)。(a-d)tt3.1突变体(a)、tt3.2突变体(b)、过表达TT3.1WYJ(c)和过表达TT3.1CG14(d)转基因水稻植株在正常条件下和42℃热胁迫7天后的表型。
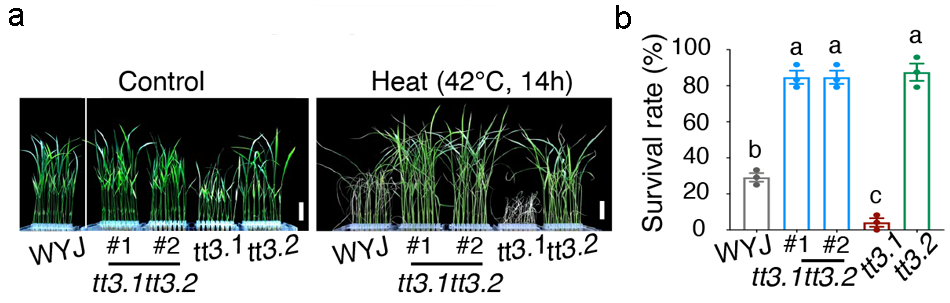
图3 tt3.1 tt3.2双突变体耐热表型分析(Zhang et al., 2022)。(a)tt3.1 tt3.2双突变体在正常条件下和42℃热胁迫7天后的表型;(b)tt3.1 tt3.2双突变体在正常条件下和42℃热胁迫7天后的存活率。
假设基因A具有负调控作用,基因B具有正调控作用。首先分别创制基因A和基因B的过表达株系,比较基因A过表达株系(OE-A)与野生型(WT)的表型差异,以确定基因A的负调控作用;比较基因B过表达株系(OE-B)与野生型(WT)的表型差异,以确定基因B的正调控作用。然后创制双过表达株系,比较双过表达株系(OE-AB)与单基因过表达株系(OE-A和OE-B)的表型差异,分析基因A和基因B之间的调控关系:
如果基因A过表达株系(OE-A)的表型是某些特定性状缺失或减弱,而双过表达株系(OE-AB)表型恢复至野生型或接近野生型表型,或者双过表达株系(OE-AB)表型与B过表达株系一致,说明基因B的过表达能够恢复基因A过表达引起的表型缺失或减弱。这种情况表明,基因A在基因B的上游。如案例3、案例4所示。
如果基因A过表达株系(OE-A)的表型是某些特定性状缺失或减弱,而双过表达株系(OE-AB)表型与A过表达株系一致,说明基因B的过表达不能恢复基因A过表达引起的表型缺失或减弱。这种情况表明,基因B在基因A的上游。但是这种情况小远未找到相应的文献案例,如果大家有相关的文献,记得分享给小远噢。
案例3
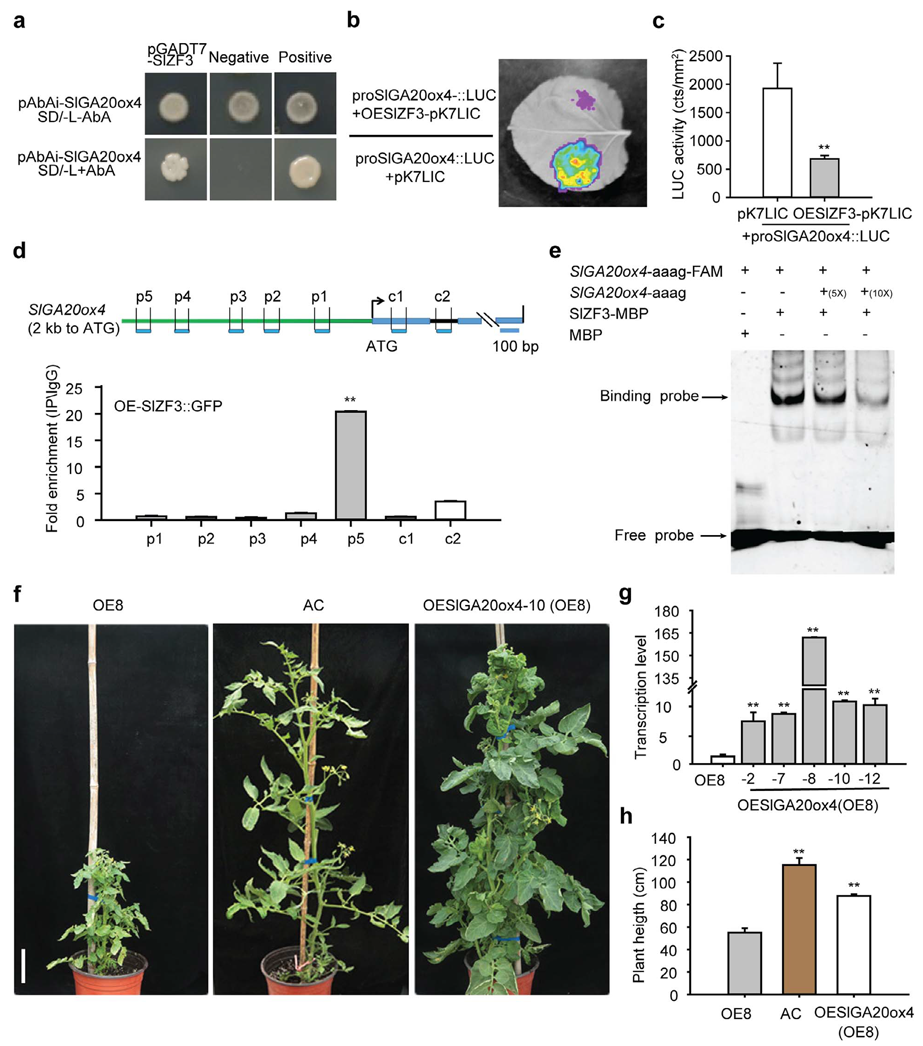
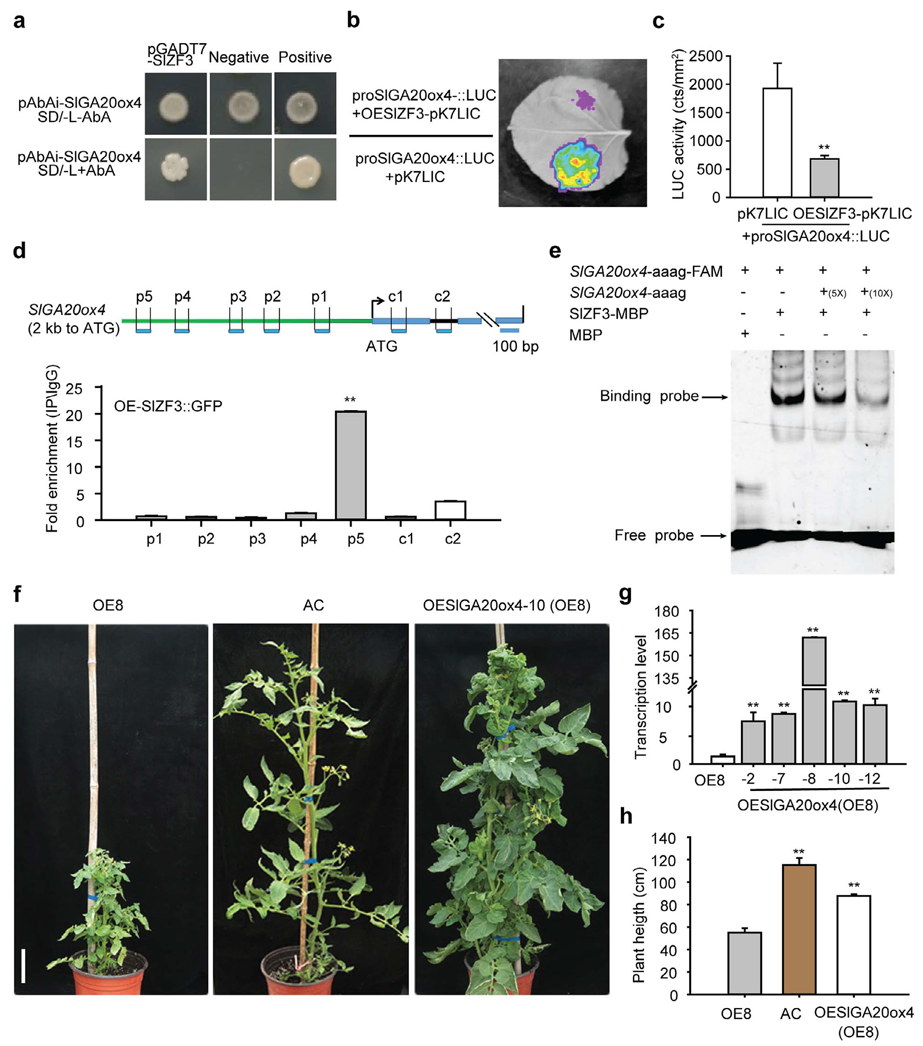
2023年4月,华中农业大学欧阳波课题组在Horticulture Research杂志上发表了一篇题为“SlZF3 regulates tomato plant height by directly repressing SlGA20ox4 in gibberellic acid biosynthesis pathway”的研究论文。作者发现SlZF3负调控番茄株高,过表达SlZF3可导致番茄矮化。随后作者利用Y1H、Dual-LUC、ChIP-qPCR和EMSA实验证明SlZF3可直接结合到SlGA20ox4的启动子上(图4a-e)。为了进一步探究SlZF3和SlGA20ox4的遗传关系,作者在SlZF3过表达株系的背景上进一步过表达SlGA20ox4,获得SlZF3 SlGA20ox4双过表达株系,发现SlZF3 SlGA20ox4双过表达株系的株高与野生型相似,说明SlZF3过表达对株高的抑制作用在SlGA20ox4过表达背景中得到缓解(图4f-h),从遗传关系上进一步揭示SlZF3在SlGA20ox4的上游发挥作用来调控株高。
案例4
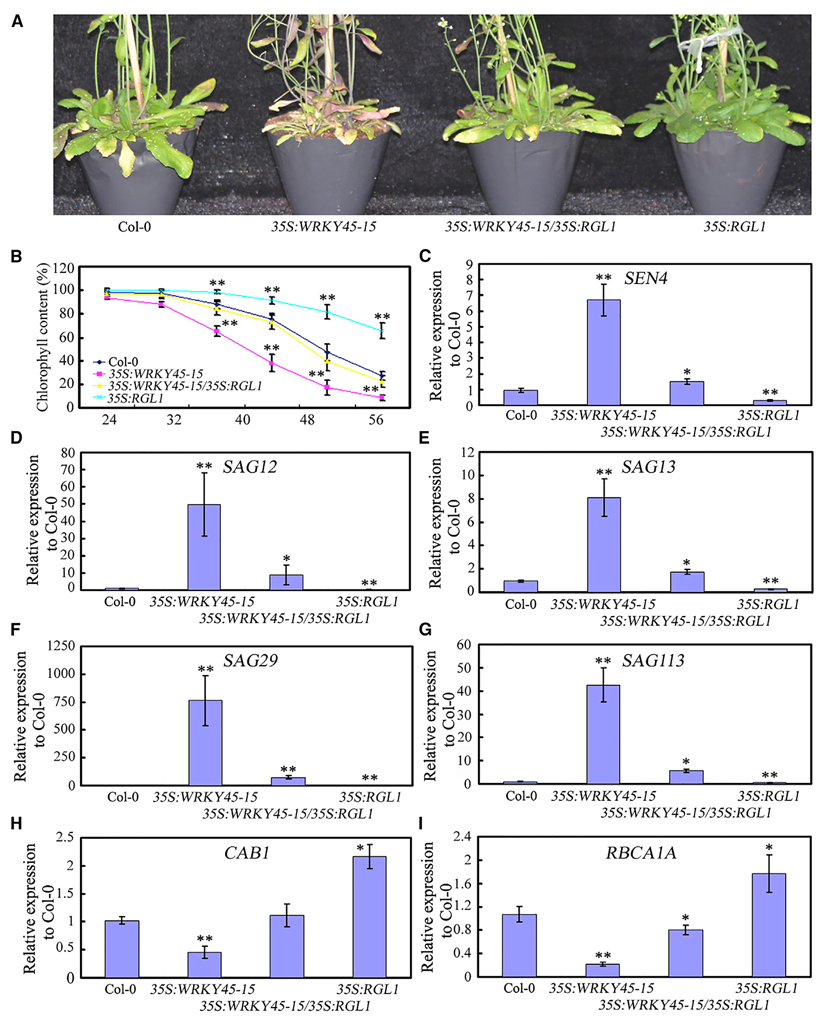
2017年9月,中国科学院西双版纳热带植物园余迪求课题组在Molecular Plant杂志上发表了一篇题为“Arabidopsis WRKY45 interacts with the DELLA protein RGL1 to positively regulate age-triggered leaf senescence”的研究论文。作者发现WRKY45是植物衰老的正调控因子,WRKY45基因突变能延缓植物衰老进程,而其过表达则显著促进植物衰老进程。进一步研究发现WRKY45能与RGL1蛋白在细胞核内相互作用形成复合物共同调控植物衰老进程。同时作者分析发现RGL1是植物衰老的负调控因子,RGL1过表达能显著延缓植物衰老。为了研究WRKY45和RGL1的遗传关系,作者将35S:RGL1与35S:WRKY45-15杂交得到35S:WRKY45-15/35S:RGL1双过表达株系,发现与35S:RGL1相比,35S:WRKY45-15/35S:RGL1双过表达株系表现出早期衰老表型,说明WRKY45的过表达能够使RGL1过表达延缓植物衰老的表型得到抑制(图5)。这些结果表明,RGL1在WRKY45的上游发挥作用从而调控植物衰老。
如果基因A突变体表现出某些特定性状缺失或减弱,而突变体A/过表达B杂交材料表型与基因A突变体表型相似,说明基因B的过表达不能补偿基因A突变引起的表型缺失或减弱。这种情况表明,基因B在基因A的上游。如案例6所示。
案例5
2021年1月,四川大学刘建全课题组在New Phytologist杂志上发表了一篇题为“WRKY33 interacts with WRKY12 protein to up-regulate RAP2.2 during submergence induced hypoxia response in Arabidopsis thaliana”的研究论文,揭示了WRKY33/WRKY12-RAP2.2分子模块调控拟南芥水淹低氧适应的新机制。作者发现WRKY33、WRKY12和RAP2.2均正向调控水淹环境中的缺氧反应,与野生型Col相比,三个基因的过表达拟南芥材料均呈现出明显的抗水淹胁迫表型,例如存活率增加,叶片损伤较小,过氧化物积累减少,MDA积累减少。为了进一步研究WRKY33、WRKY12和RAP2.2在水淹低氧途径中的遗传关系,作者通过遗传杂交,获得wrky12 RAP2.2-OE和wrky33 RAP2.2-OE拟南芥材料并进行水淹表型分析,发现过表达RAP2.2能分别恢复wrky12和wrky33突变体的水淹胁迫表型(图6)。表明WRKY12和WRKY33转录因子可能通过RAP2.2正向调控缺氧应激反应,WRKY33和WRKY12可能在缺氧反应途径中位于RAP2.2的上游。
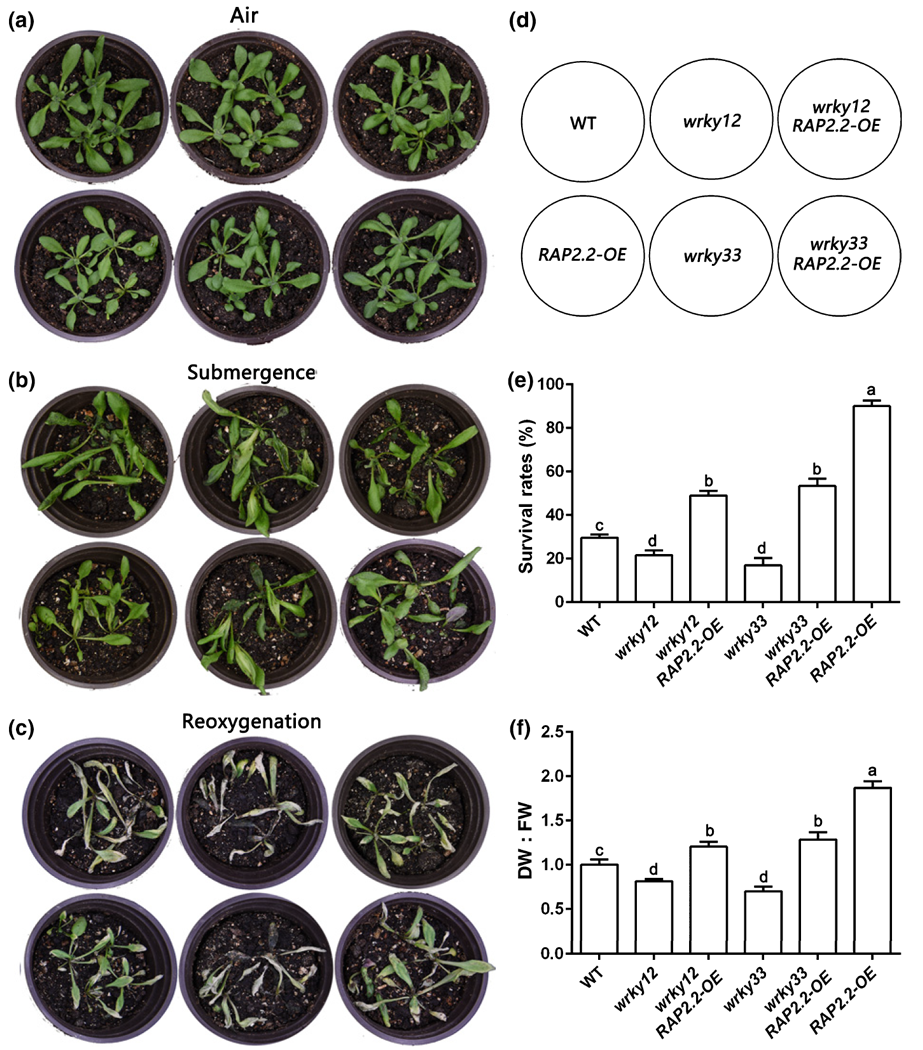
图6 WRKY12、WRKY33和RAP2.2之间的遗传关系分析(Tang et al., 2021)。(a-c)4周龄的拟南芥野生型(WT)、wrky12、wrky33、RAP2.2-OE、wrky12 RAP2.2-OE和wrky33 RAP2.2-OE植物的表型:(a)淹水前、(b)淹水48小时后、(c)恢复4天后;(d)相应植物的示意图;(e)植株的存活率;(f)恢复4天后植株的DW : FW比率。FW为初始鲜重,DW为干重;WT的DW : FW比率设为1。
案例6
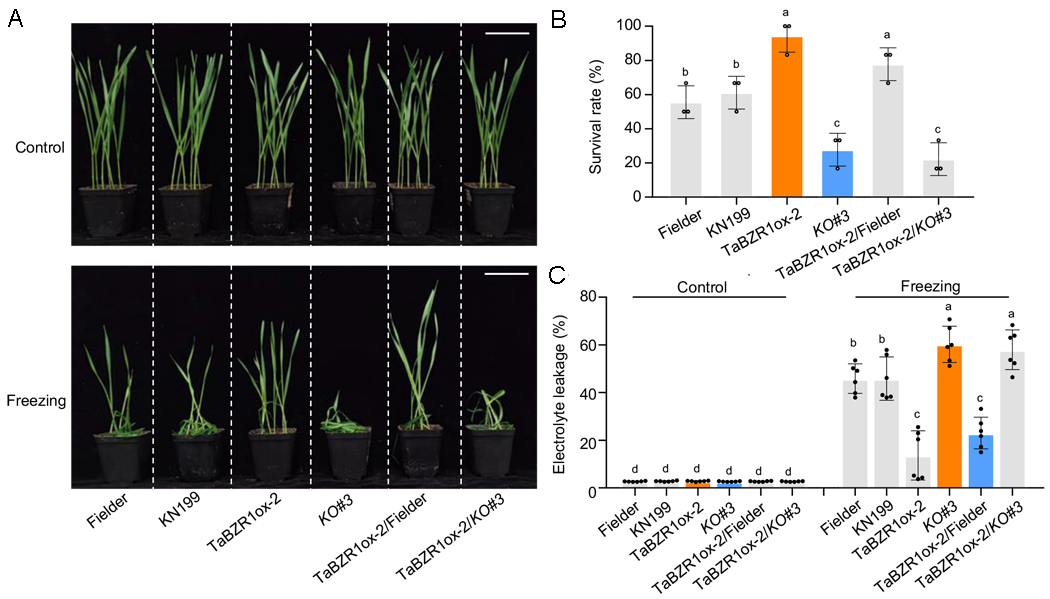
2024年7月,中国农业大学胡兆荣课题组在The Plant Cell杂志上发表了一篇题为“Methyltransferase TaSAMT1 mediates wheat freezing tolerance by integrating brassinosteroid and salicylic acid signaling”的研究论文。作者发现TaSAMT1可能通过影响MeSA合成正向调控小麦耐冷性,与野生型相比,TaSAMT1-OE株系耐冷性显著提升,而敲除株系耐冷性则显著降低。随后作者筛选鉴定到TaSAMT1基因的上游调控蛋白TaBZR1,并通过超表达株系和突变体表型鉴定发现TaBZR1能够正向调控小麦耐冷性。为了进一步确定TaSAMT1和TaBZR1在小麦耐冷性调控中的遗传关系,作者将TaBZR1过表达系(ox-2)与Tasamt1突变体(KO#3)杂交,发现TaBZR1ox-2/Tasamt1 KO#3株系与Tasamt1突变体表型相似,均表现出较低的耐冷性,说明TaBZR1的过表达不能恢复TaSAMT1突变导致的耐冷性降低(图7),表明TaBZR1在TaSAMT1的上游起作用并参与小麦耐冷性调控。
如果基因A过表达表现出某些特定性状缺失或减弱,而过表达A/突变体B杂交材料表型恢复至野生型或接近野生型表型,说明基因B的敲除能够恢复基因A过表达导致的表型缺失或减弱。这种情况表明,基因A在基因B的上游。如案例8所示。
案例7
2024年1月,南京农业大学徐国华课题组、顾冕课题组和上海市农业生物基因中心严明课题组联合在The Plant Cell 杂志上发表了一篇题为“The transcription factor MYB110 regulates plant height, lodging resistance, and grain yield in rice”的研究论文。作者发现MYB110 是株高的负调控因子,OsMYB110 突变后使水稻株高显著增加。PHR2也是株高的负调控因子,OsPHR2过表达植株的株高均低于WT植株。此外,作者发现OsPHR2可直接与MYB110 启动子中的P1BS基序结合。为了研究OsPHR2和MYB110 之间潜在的遗传相互作用,作者进行了PHR2-Ox株系和myb110 突变体之间的杂交,发现MYB110 在PHR2-Ox背景中的突变使株高恢复到与myb110 突变体相当的水平(图8A、B)。相比之下,MYB110-Ox背景中OsPHR2的突变并不能挽救MYB110-Ox植物的迟缓生长(图8C、D)。同时,myb110 phr2双突变体的高度与myb110 植株相似(图8E、F)。这些结果表明,OsPHR2在MYB110 上游发挥作用调节植物株高。
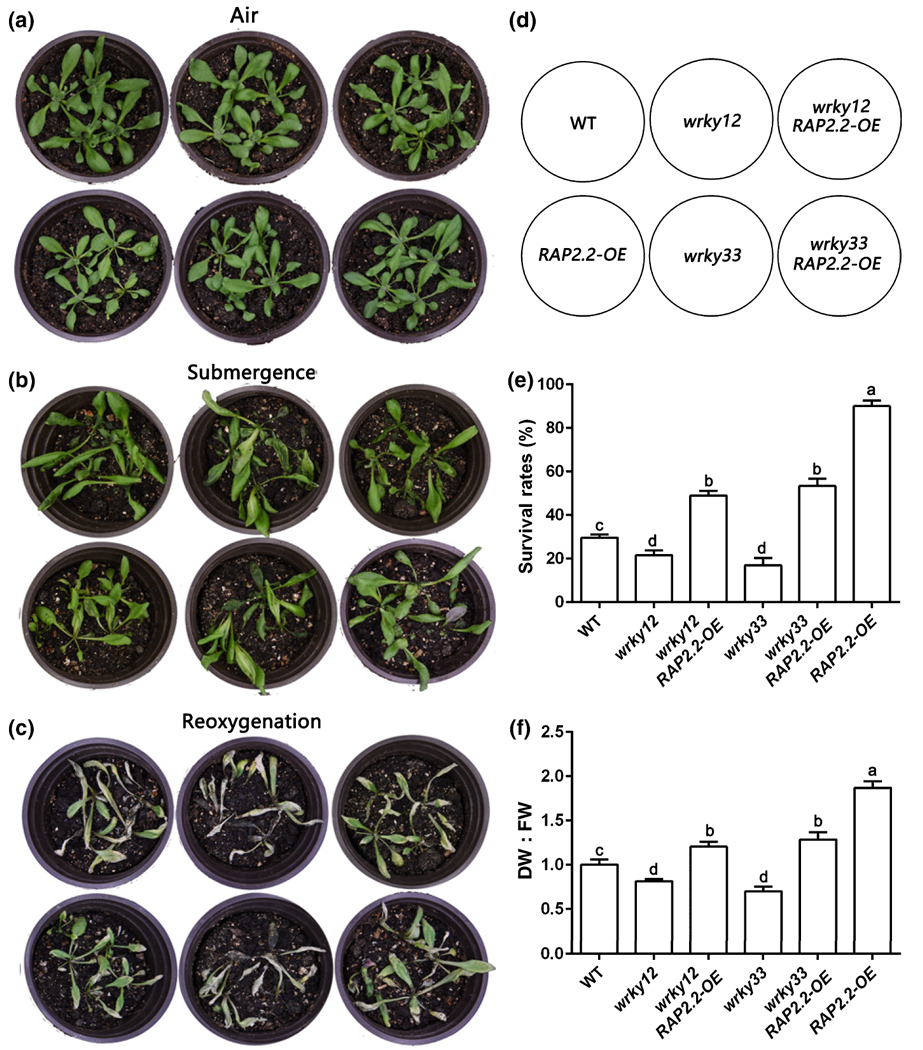
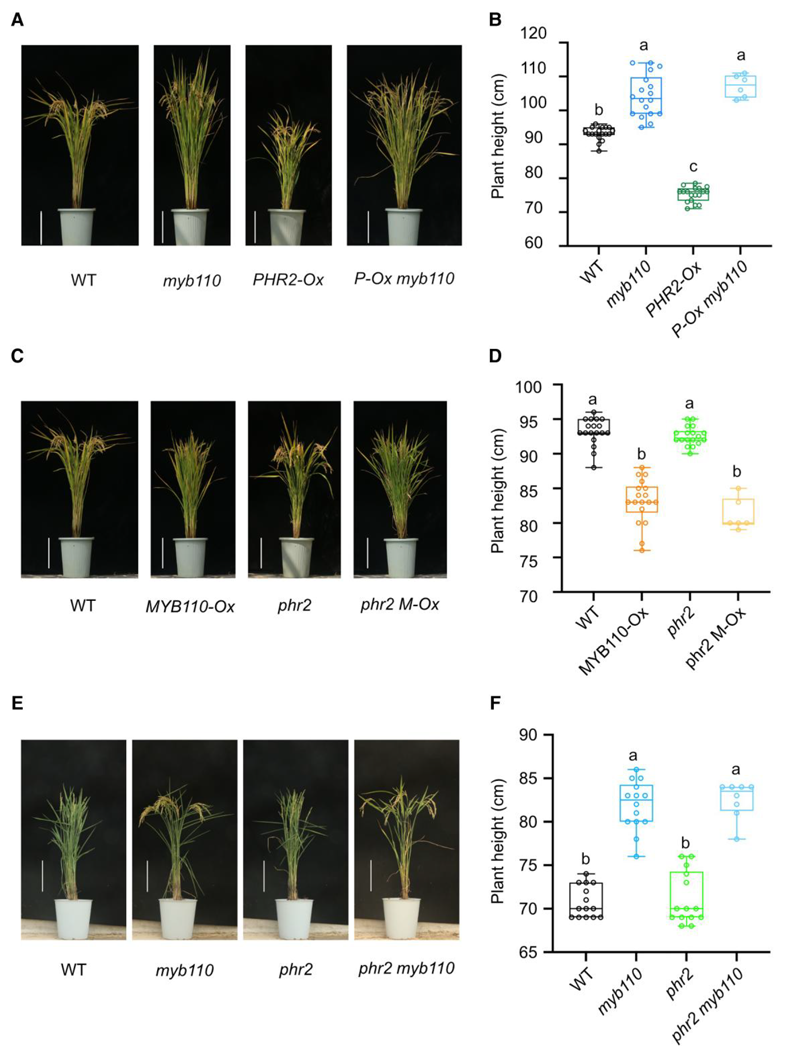
图8 PHR2在遗传上作用于MYB110上游并负调控植株高度(Wang et al., 2024)。(A、B)野生型(WT)、myb110、PHR2-Ox及其杂交材料植株表型(A)和植株高度(B);(C、D)野生型(WT)、MYB110-Ox、phr2及其杂交材料植株表型(C)和植株高度(D);(E、F)野生型(WT)、myb110、phr2及其杂交材料植株表型(E)和植株高度(F)。
案例8
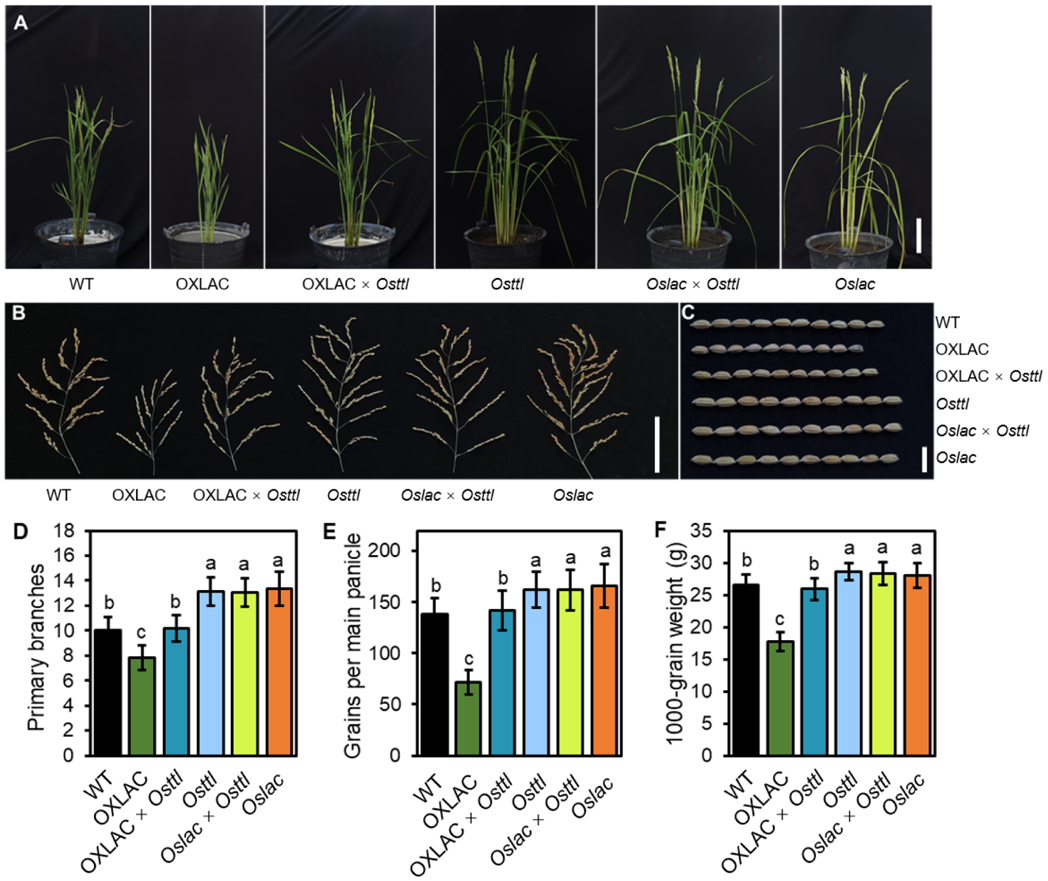
2024年3月,中山大学陈月琴/练剑平课题组和广东省农科院于洋课题组联合在The Plant Cell杂志上发表了一篇题为“A transthyretin-like protein acts downstream of miR397 and LACCASE to regulate grain yield in rice”的研究论文。在前期研究中,作者发现漆酶基因OsLAC负调控植株高度和籽粒大小,但其作用机制尚不清楚(Zhang et al., 2013)。在该研究中,作者对OsLAC进行了生化和遗传学分析,发现OsTTL蛋白可与OsLAC相互作用。同时发现OsTTL负调控植株高度、籽粒大小和产量,在水稻中过表达OsTTL导致植株呈现株型矮小、叶片直立、籽粒变小和产量降低等表型,而敲除OsTTL基因则导致植株株型变大、籽粒增大和产量提高。为了进一步研究OsLAC和OsTTL之间潜在的遗传关系,作者将Osttl突变体与OsLAC过表达(OXLAC×Osttl)或敲除(Oslac×Osttl)植株进行杂交,发现OXLAC×Osttl株系的表型与野生型相似,说明OsTTL的敲除恢复了OXLAC植株的发育缺陷(OXLAC×Osttl),包括植物结构、穗分枝和籽粒大小的缺陷。Oslac×Osttl双突变体的产量相关性状与Osttl植株相似(图9)。这些结果表明OsLAC在OsTTL上游起作用以调节植株高度和籽粒产量。