本文内容速览:
对自交不亲和植物而言,其与自交亲和植物在空间、时间和传粉方式上均存在不同。在空间上,自交不亲和植物的雄花和雌花分别出现在不同的植株上或不同的花序上以避免自交。在时间上,自交不亲和植物的雄花和雌花在不同的时间点成熟,从而减少自交的可能性。在传粉方式上,许多自交不亲和植物依赖动物媒介进行传粉,通过吸引不同的传粉者,以降低自交的频率。
在自交不亲和过程中,涉及植物的雌蕊对“自已”和“异已”花粉的识别,这种识别通常是由单个高度多态性遗传位点S位点(在某些科中是两个或多个S位点)控制的,S位点包含了紧密连锁的雌蕊决定因子和花粉决定因子,整个S位点是一个完整的遗传单元,被称为S单体型。当雌蕊-花粉相互识别时,如果雌蕊和花粉具有相同的S单体型,会迅速启动一系列的信号转导,导致雌性器官不接受自已的花粉,反之,如果雌蕊和花粉具有不同的S单体型,则受精过程正常进行。
表1 不同植物的自交不亲和类型(信息来自Zhang et al., 2024)。
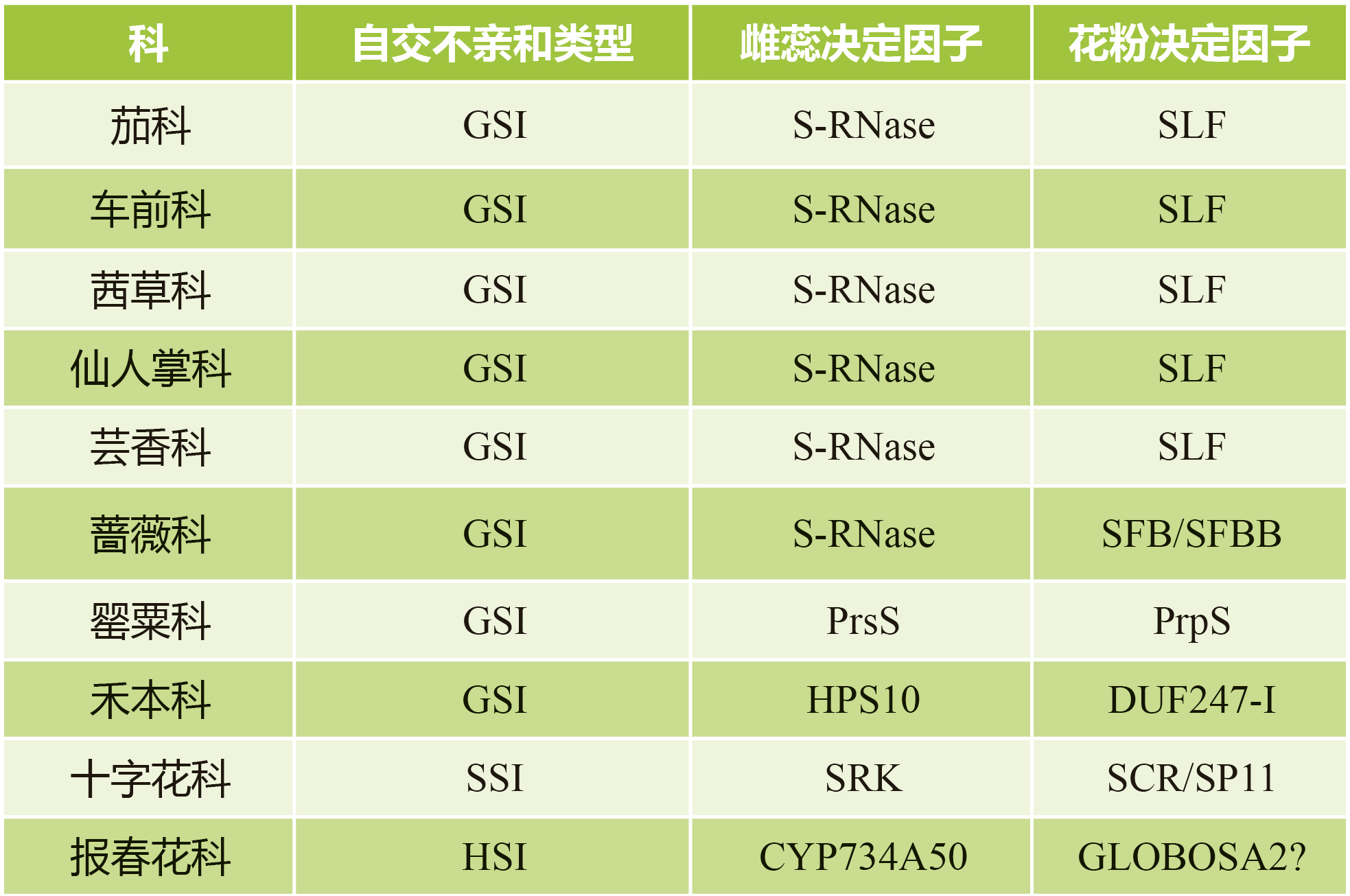
备注:表中所列为该科代表性物种的自交不亲和机制,每个科中仍可能存在差异,具体的物种具体分析。“?”指目前的研究还不是十分确定为该因子。
现在咱们来了解一下同型自交不亲和中的GSI和SSI(图1)。GSI的花粉亲和与否,由单倍体花粉(即配子体)自身的S基因型决定,与亲本的基因型无关,胚珠和花粉为相同的S单体型时会表现出不亲和,其大部分表现为花粉可以正常的水合、萌发及花粉管穿过柱头,但在花柱中,花粉管发生破裂因而不能到达胚珠。而SSI的花粉亲和与否的表型由产生花粉的二倍体亲本(即孢子体)的S基因型决定,与配子体的基因型无关,SSI反应主要发生在柱头表面,表现为花粉落到柱头上引发柱头表面产生胼胝质以阻碍花粉穿过柱头乳突细胞。
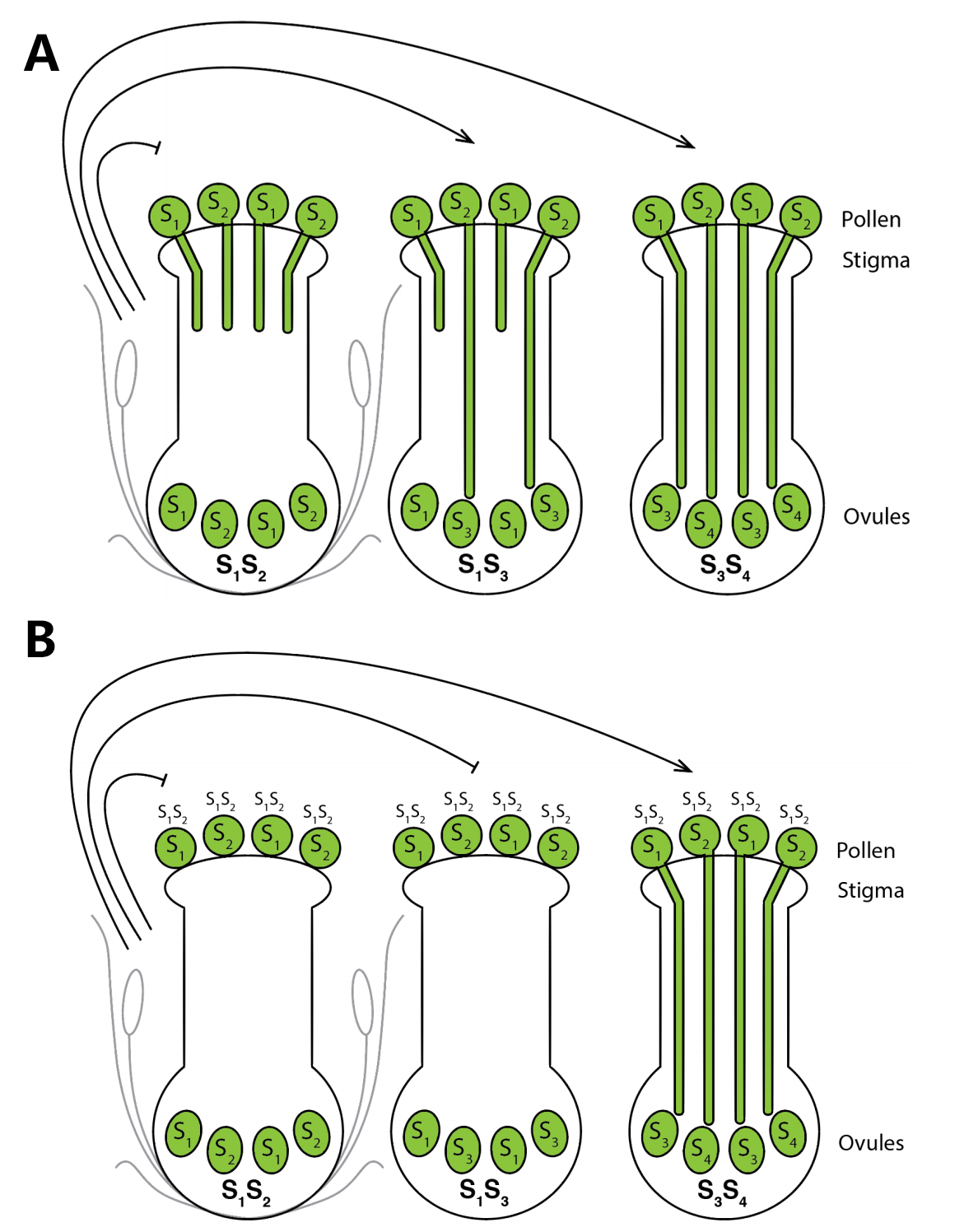
图1 GSI和SSI的模式图(Bachmann JA, 2020)。亲本的S位点基因型在图中用SnSn表示。(A)在GSI系统中,只有雌雄配子的基因型一致时,才会产生自交不亲和的表型:左侧为不亲和自花授粉,花粉管生长受阻,右侧为不同S单体型个体之间的异花授粉,中间因其有一个不同的S单体型因而发生部分亲和,导致花粉管发育阻滞;(B)在SSI系统中,由花药绒毡层和柱头乳突细胞的基因型决定自交不亲和的表型:左侧为不亲和自花授粉,右侧为不同S单体型个体之间的异花授粉,中间因其有一个相同的S单体型因而导致不亲和,花粉不萌发。
在长期进化中,不同物种形成了不同的自交不亲和反应途径和机制,其中有一些被研究得较多,如下图所示。
GSI系统是最常见的SI机制,GSI主要分为S-核酸酶系统(S-RNase)和SI-细胞程序性死亡(SI-PCD)系统。
茄科、车前科、茜草科、仙人掌科、芸香科和蔷薇科是由雌蕊决定因子S-RNase介导的GSI系统(表1)。在亲和授粉中,S-RNase被降解和失活,花粉管得以生长,在不亲和授粉中,S-RNase被激活,参与花粉管生长的RNA被降解。
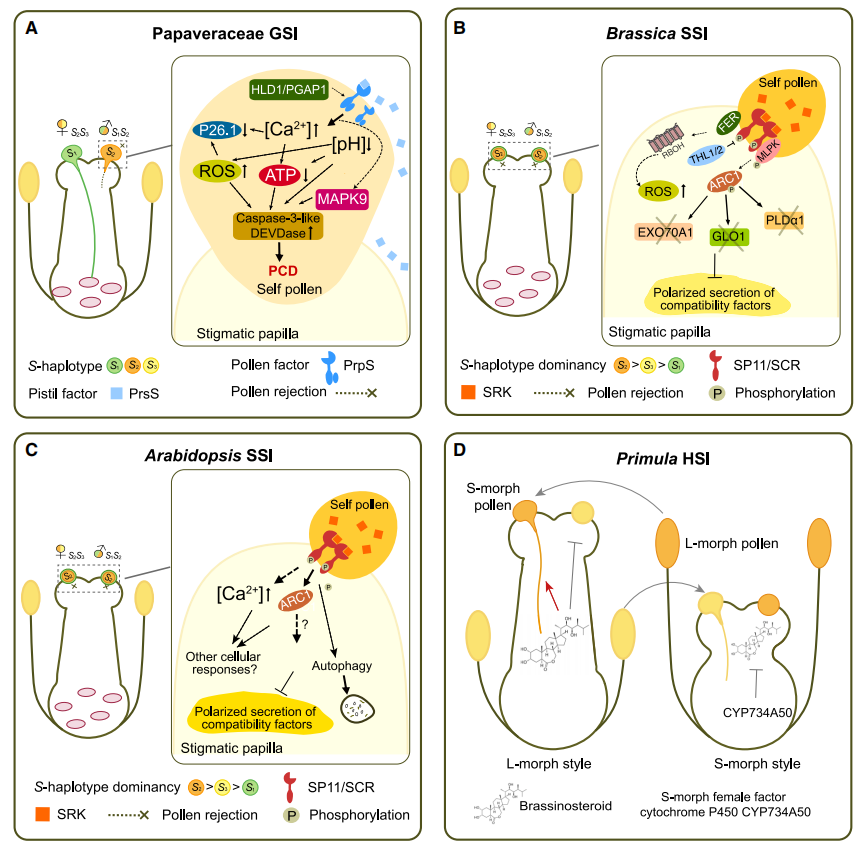
在罂粟科中是以SI-PCD介导的GSI机制,不同于上述的GSI,其雌蕊决定因子是一种由柱头乳突细胞分泌到雌蕊表面的蛋白质PrsS,而花粉决定因子是一种位于花粉中的跨膜蛋白PrpS,在不亲和授粉中,当雌蕊-花粉相互识别后,导致细胞内Ca2+和活性氧(ROS)的增加,诱导多层信号级联反应,最终导致自花花粉即不亲和花粉的PCD(图2A)。
禾本科GSI系统不同于上述GSI系统,其由两个互不连锁的多等位基因位点S和Z控制,S位点表达一个雌蕊决定因子sS和两个花粉决定因子SDUF247-I和SDUF247-II,Z位点表达一个雌蕊决定因子sZ和两个花粉决定因子ZDUF247-I和ZDUF247-II,当雌蕊的S基因型与花粉的S基因型相同,以及雌蕊的Z基因型与花粉的Z基因型相同时,植物会表现出自交不亲和。
除了上述介绍的三种GSI之外,十字花科中的芸薹属、拟南芥属的自交不亲和现象属于SSI,雌蕊决定因子为受体激酶SRK,花粉决定因子为SCR小肽(也被称为SP11)。在不亲和授粉中,当雌蕊-花粉相互识别后,SRK和SCR/SP11相互作用并在柱头乳突细胞中发生一系列的信号级联反应,导致对自花花粉的排斥(图2B、C)。
前面也提到,除了同型自交不亲和GSI和SSI之外,还存在异型自交不亲和,以报春花科HSI为例,其表现为雌蕊和花粉在空间上的分离,报春花的花柱形态和其SI是连锁的,都由细胞色素P450 CYP734A50决定,CYP734A50的表达可抑制花柱中油菜素内酯的活性,抑制细胞的伸长以形成短花柱(图2D)。
但是,当科学家们在利用自交不亲和系选育F1代杂交种时,也无法避免F1代的亲本自交不亲和程度高导致其自身繁种困难这个难题。针对这个现象,可以使用物理、化学等方法处理柱头以克服自交不亲和反应,或者,干扰或敲除自交不亲和相关基因以获得自交亲和系(图3)。
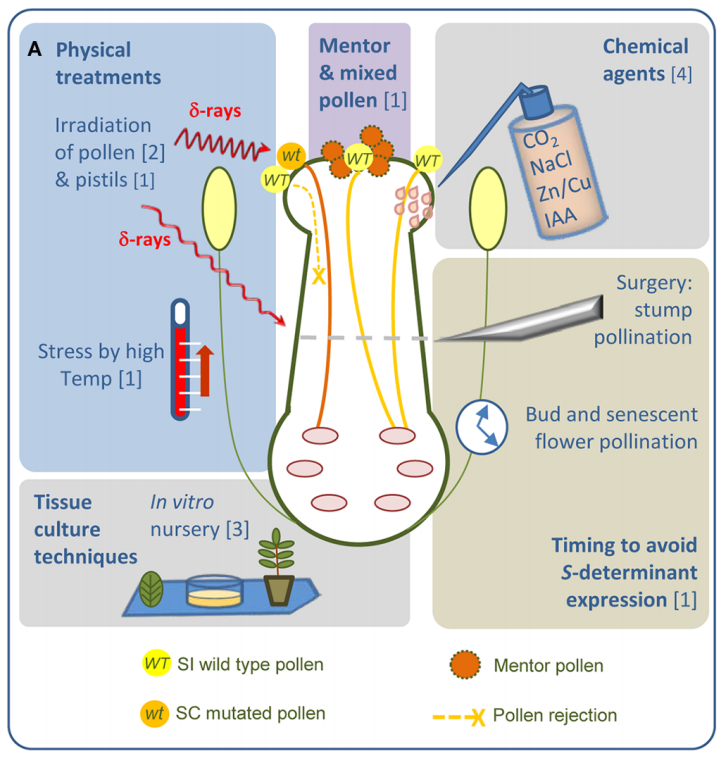
2018年8月,中国农科院黄三文和张春芝课题组在Nature Plants杂志上发表了一篇题为“Generation of self-compatible diploid potato by knockout of S-RNase”的研究论文,作者通过CRISPR/Cas9基因编辑技术敲除马铃薯S-RNase基因,使S-RNase蛋白失活,成功创制自交亲和的二倍体马铃薯材料,为马铃薯的品种改良展现了巨大的前景。

2021年4月,华东师范大学李超课题组在Science杂志上发表了一篇题为“PCP-B peptides unlock a stigma peptide-receptor kinase gating mechanism for pollination”的研究论文,作者以十字花科拟南芥为研究对象,揭示了花粉通过其覆盖物中的PCP-Bγ小肽竞争柱头中的RALF33小肽,进而抑制柱头中RALF33-FER/ANJ受体激酶信号通路维持的ROS水平,从而影响花粉水合和萌发的柱头-花粉识别的分子机理。
2023年1月,山东农业大学段巧红和马萨诸塞大学Alice Y. Cheung课题组合作在Nature杂志上发表了一篇题为“Stigma receptors control intraspecies and interspecies barriers in Brassicaceae”的研究论文,作者揭示了十字花科蔬菜通过调控柱头ROS水平以维持种间生殖隔离的分子机制,并通过外源施加ROS清除剂(Na-SA)、NO供体(GSNO)或使用反义寡核苷酸(AS-ODN)技术成功获得大白菜种间、属间远缘杂交胚,为创制新种质提供了重要思路。
2023年10月,北京大学瞿礼嘉和钟声课题组在Cell杂志上发表了一篇题为“Antagonistic RALF peptides control an intergeneric hybridization barrier on Brassicaceae stigmas”的研究论文,作者在分子水平上解析了拟南芥柱头识别并接受同种花粉而不接受远缘花粉的机制,提出了柱头-花粉间识别与信号交流的“锁-钥模型”,阐明了柱头处的种间、属间生殖障碍形成的机理,完美解释了“花粉蒙导效应”,并可通过施加人工合成的pRALF小肽即“钥匙”到柱头上,即可打开柱头处的“锁”,让远缘花粉穿入柱头以克服生殖障碍,并最终获得了不同属间的远缘杂交胚。
Bachmann JA (2020) Evolutionary consequences of dominance at the Brassicaceae self- incompatibility locus.
Boris I, Russell L, Kohn JR (2008) Loss of Self‐Incompatibility and Its Evolutionary. INT J PLANT SCI 169: 93-104
Dou S, Zhang T, Tu J, Shen J, Yi B, Wen J, Fu T, Dai C, Ma C (2021) Generation of novel self-incompatible Brassica napus by CRISPR/Cas9. PLANT BIOTECHNOL J 19: 875-877
Muñoz-Sanz JV, Zuriaga E, Cruz-García F, McClure B, Romero C (2020) Self-(In)compatibility Systems: Target Traits for Crop-Production, Plant Breeding, and Biotechnology. FRONT PLANT SCI 11
Ye M, Peng Z, Tang D, Yang Z, Li D, Xu Y, Zhang C, Huang S (2018) Generation of self-compatible diploid potato by knockout of S-RNase. NAT PLANTS 4: 651-654
Zhang D, Li YY, Zhao X, Zhang C, Liu DK, Lan S, Yin W, Liu ZJ (2024) Molecular insights into self-incompatibility systems: From evolution to breeding. PLANT COMMUN 5: 100719
