磷酸化磷脂酰肌醇 (PtdIns)是生物膜甘油磷脂的成分之一,其D-肌醇环的3、4和5位羟基可发生多重磷酸化反应形成多样的衍生物,共同构成复杂的磷脂酰肌醇代谢网络。其中,3,5-二磷酸磷脂酰肌醇(PtdIns (3,5) P2)在内膜运输、信号转导、细胞极性生长以及细胞骨架重组等多个细胞发育过程中具有关键调控功能。FAB1 (Formation of aploid and binucleate cells 1)/PIKfyve (FYVE finger-containing phosphoinositide kinase)激酶催化PtdIns3P是PtdIns (3,5) P2合成的唯一途径。拟南芥具有四个 FAB1 同源物(FAB1A 至 D),其中 FAB1A 和 FAB1B 是典型的,具有识别所需的 FYVE 结构域。相比之下,FAB1C 和 FAB1D 缺乏这种典型的 FYVE 结构域。
有研究表明,FAB1 在跨膜运输中发挥作用,包括 PIN2。但FAB1如何影响真核生物PIN运输仍不清楚。近日,Kwang-Ho Maeng等人在国际著名学术期刊PNAS发表了题为“FAB1C, a phosphatidylinositol 3-phosphate 5-kinase, interacts with PIN-FORMEDs and modulates their lytic trafficking in Arabidopsis”的文章,该文主要研究发现FAB1C与生长素输出载体互作并参与PIN的质膜运输。
生长素是具有核心作用的激素,控制着植物生长和发育的几乎所有方面。其输出载体主要由PIN(PIN-FORMED)家族组成,包括8个成员(PIN1-8),控制生长素从细胞膜向细胞外空间的输出。PIN蛋白(PIN1、PIN2、PIN3、PIN4 和 PIN7)结构上中间含有一个被五个跨膜螺旋保卫的亲水环(Hydrophilic loop, HL),HL结构域的翻译后修饰,如磷酸化、泛素化等,可以影响PIN蛋白的跨膜运输。
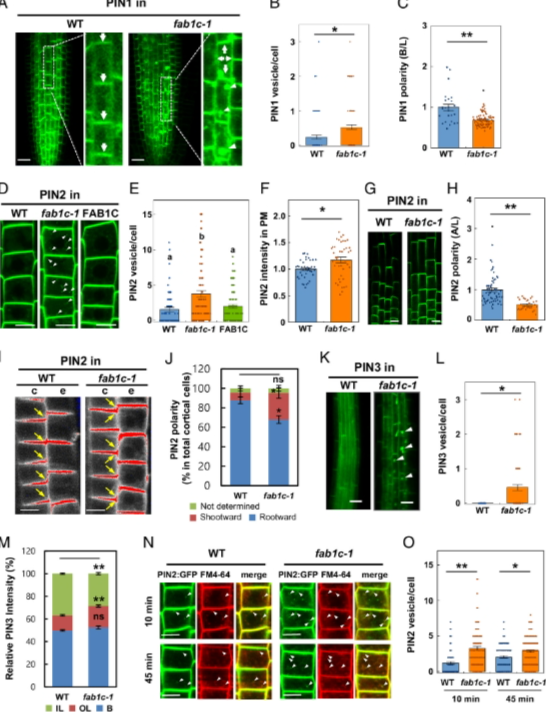
该团队研究发现FAB1C直接与 PIN-HL 相互作用并调节 PIN蛋白的跨膜运输(图1)。FAB1C 功能丧失会改变 PIN 的运输和极性(图2)。
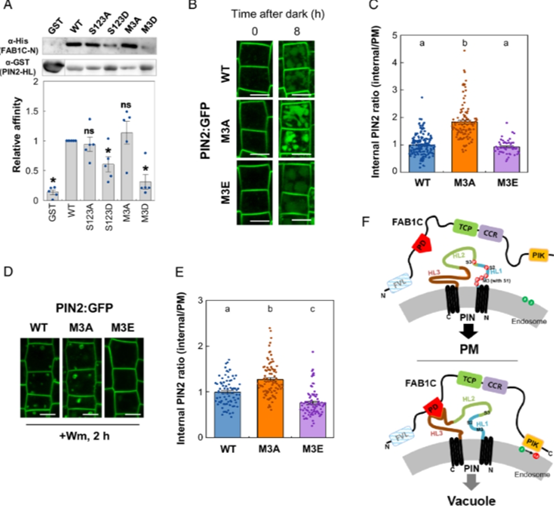
该团队研究发现,未磷酸化的 PIN2 比磷酸化的 PIN2 更容易发生液泡运输(图3)。