细胞壁决定了植物细胞的形状和功能。原生细胞壁中的纤维素微纤维通过细胞内的膨压限制细胞膨胀,从而导致细胞垂直于微纤维定向生长。纤维素微纤维沉积的模式在每种细胞类型中都有明确的确定,这使得不同组织中的细胞能够正确成形。皮层微管通过引导纤维素合酶复合物的轨迹来决定细胞壁沉积模式,纤维素合酶复合体在细胞表面合成纤维素微纤维。许多微管相关蛋白(MAPs)可以调节皮层微管的排列,例如,MOR1,RIC1和CLASP等。尽管MIDD1及其类似物已被证实参与调节皮层维管的排列,但在原生木质部细胞分化过程中,决定皮质微管排列的分子机制尚未被完全理解。近日,国际著名期刊Nature Plants在线发表了一篇题为“Microtubule-associated phase separation of MIDD1 tunes cell wall spacing in xylem vessels in Arabidopsis thaliana”的文章。该文作者通过关注原生木质部导管中的MIDD1途径,详细研究了微管排列的机制。结果表明支架蛋白MIDD1通过与超木质部导管中的ROP GTP酶和KINESIN-13A相互作用促进微管耗竭,MIDD1的微管相关相分离促进了微管排列,以调节次级细胞壁中间隙的大小。这项研究揭示了相分离在细胞壁模式微调中的一种新的生物学作用。
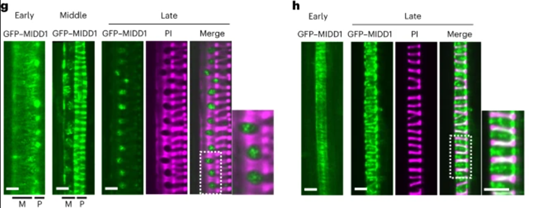
作者首先分析了MIDD1和MIDD2突变体对次生细胞壁的影响,结果发现midd1和midd2影响次生细胞壁的凹坑面积以及原生木质部导管的条纹密度。并且,条纹的密度是由皮层微管排列的改变引起的,而不是由假定的细胞生长速度较慢引起的。考虑到MIDD1与KINESIN-13A相互作用,并且MIDD1的表型与kinesin-13a的表型相当,因此,作者推测MIDD1可能通过与KINESIN-13a相互作用来调节凹坑和细胞壁间隙的大小(图1)。
图1 MIDD1调节次生细胞壁间隙的大小
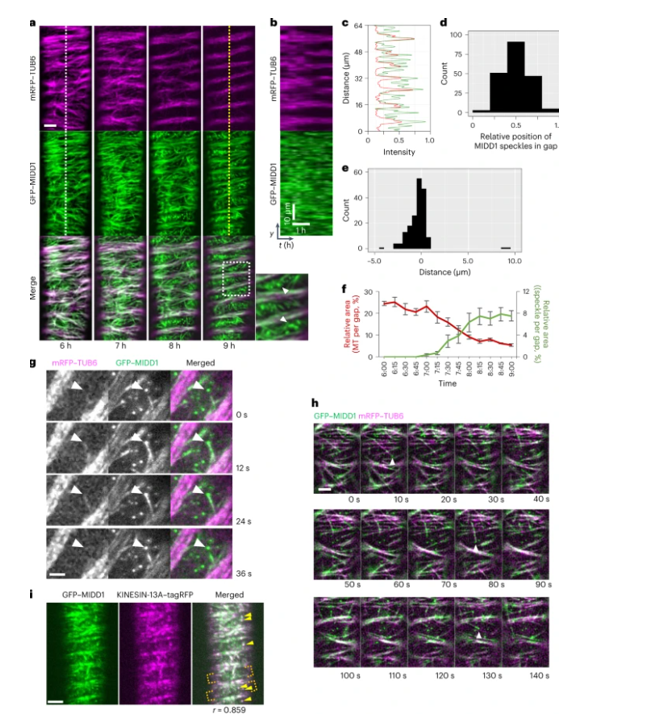
MIDD1通过ROP的活性形式被募集到质膜。作者发现ROP11和ROP7是原生木质部细胞细胞壁带密度的主要调节因子。然而,由于木质部细胞的尺寸较小位置较深,无法详细分析MIDD1和微管的。为了克服这个问题,作者通过VND7异位表达在体外诱导原木质部导管细胞分化,结果发现微管排列在midd突变体中被减弱,在分化的早期阶段,mid1斑点促进了细胞壁间隙区域的微管损耗(图2)。
图2 mid1斑点诱导的VND7GR细胞微管突变。
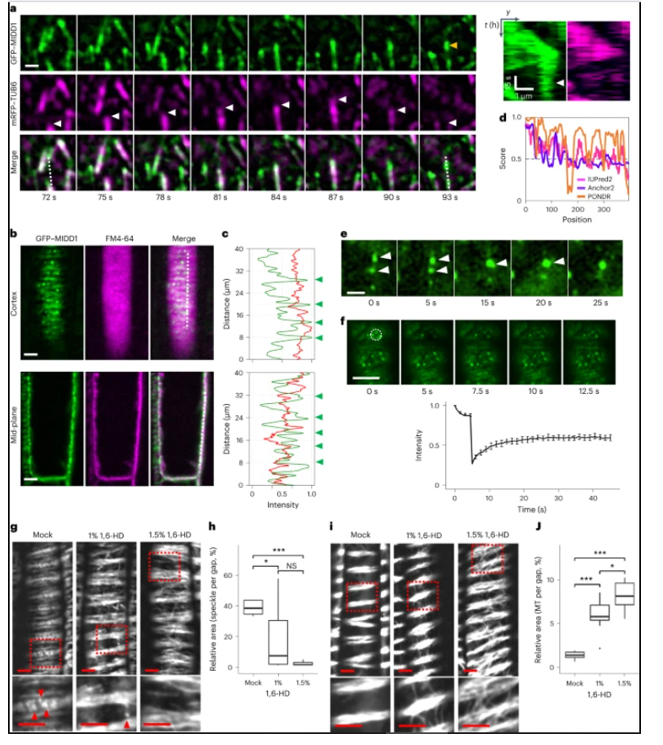
随后,作者发现微管收缩产生新的MIDD1斑点,并且新出现的MIDD1斑点沿着收缩的微管的轨迹出现。通过这些观察结果推断,MIDD1斑点不是内膜小泡,而是通过MIDD1的液-液相分离(LLPS)形成的蛋白质液滴(液体状冷凝物)。大量的MIDD1液滴反过来诱导微管的大规模解聚,从而调节微管之间的间隙大小。MIDD1液滴也在分化木质部导管中发现,这表明MIDD1相分离是原生木质部和后生木质部导管细胞壁间距的常见机制(图3)。MIDD1缩合物表现出快速周转,并且对1,6-己二醇敏感,ROP的损失消除了MIDD1的凝结,并导致原木质部导管中细胞壁间隙变窄。这一结果表明ROP直接在间隙区域形成MIDD1斑点,并且MIDD1将KINESIN-13A结合到斑点中。
图3 微管收缩触发mid1凝结
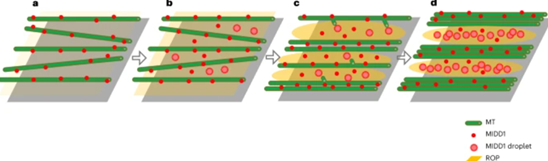
最后作者提出ROPs通过诱导微管间隙中MIDD1–KINESIN-13A斑点的形成来促进微管的分解,这种调节途径促进微管中的间隙形成,以微调细胞壁带的密度。