本文内容速览:
大家发现了嘛,科研圈里发表“背靠背”论文的现象越来越频繁了。
什么是背靠背论文呢?背靠背在英文里写为Companion papers或Back to back papers。有三种情况可发表为背靠背论文:①同一团队在同一期刊同时投稿并发表多篇关联性论文;②不同团队在互不知晓对方研究的情况下,恰好同时在同一期刊投稿,并由编辑主动撮合为背靠背论文;③不同团队在互不知晓对方研究的情况下,经过交流得知对方进展后相约一起投稿并发表。
背靠背论文为什么出现的频率越来越高了?随着技术的进步以及科研效率的提升,热点领域的研究门槛降低,科研竞争日益加剧,多个团队恰巧研究同一课题的情况逐渐增多,研究者是否能“首发”成为获得更多科研资源的重要路径之一。通过学术交流活动知晓竞争者的研究进度后进行“抢发”破坏了交流的本意,因而不同团队选择合作,并以背靠背的形式发布研究成果在一定程度上解决了恶性竞争的问题。同时,同一个结论由不同团队以不同的视角验证得来,互相补充、互相促进,更能提高领域内的影响力、获得更多的关注,成就一段佳话,共建良好学术生态。
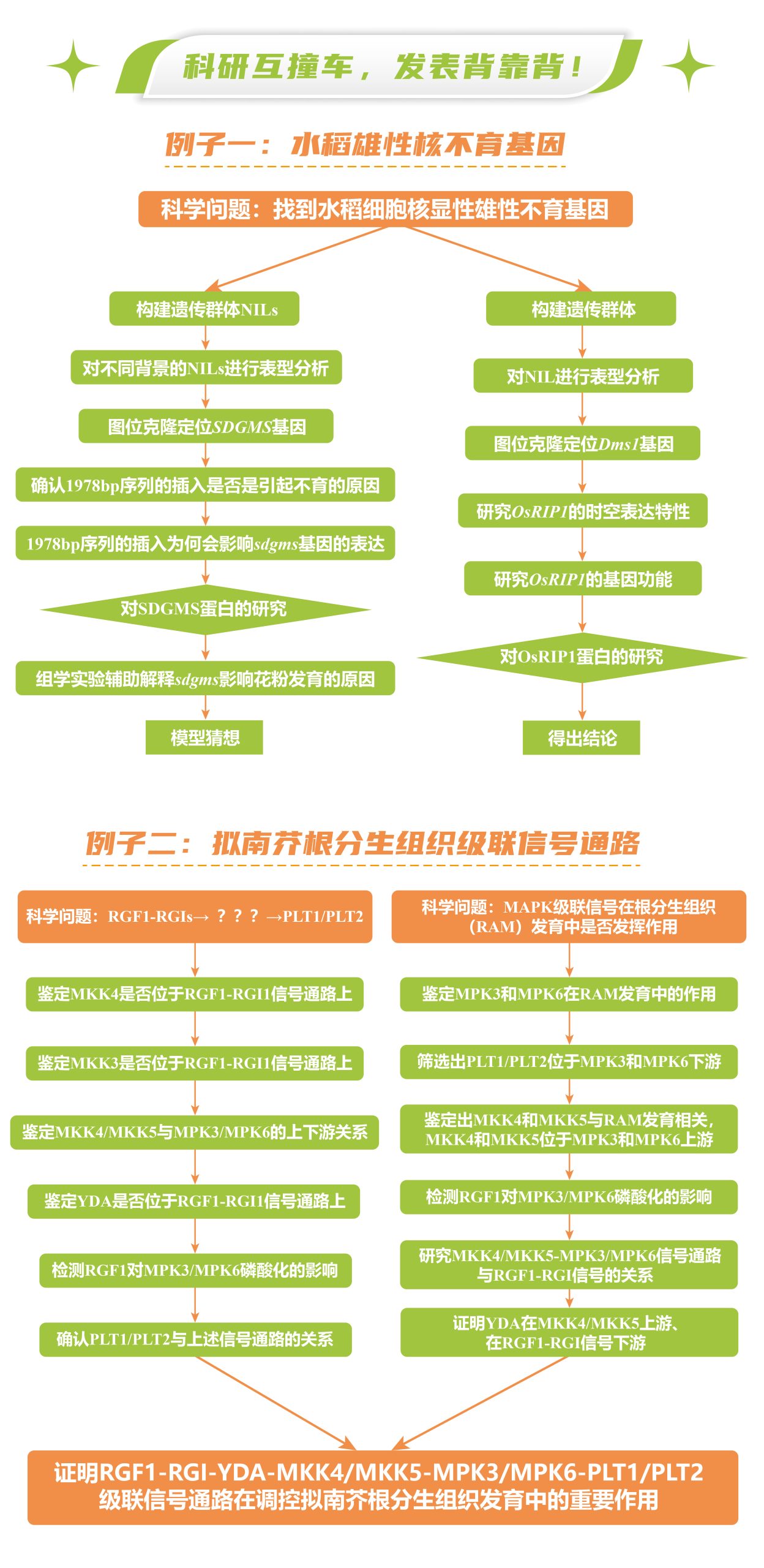
伯小远在本文列举两个植物科研圈的研究“撞车”事件,大家可以从中学习到相同的假设如何用不同的方法进行证明,以及,如何从不同的视角切入相同的研究,后续又是怎么进行研究的。
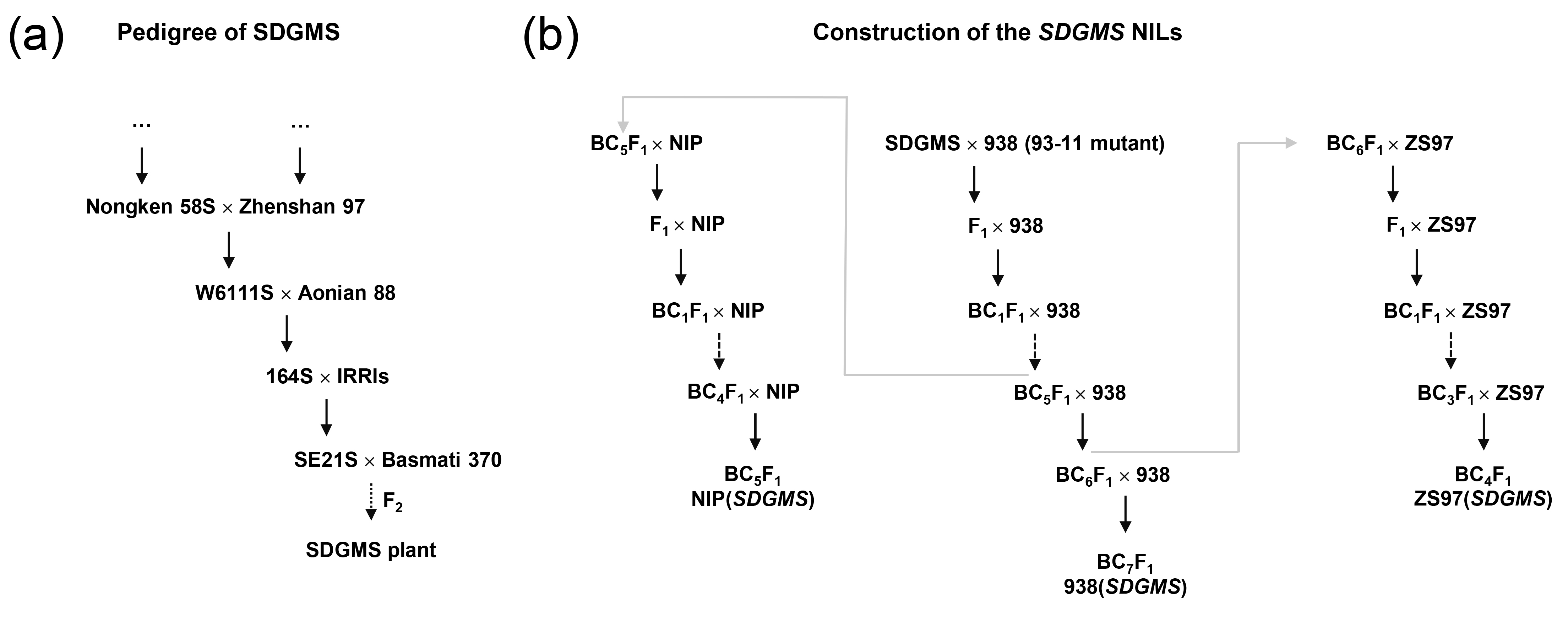
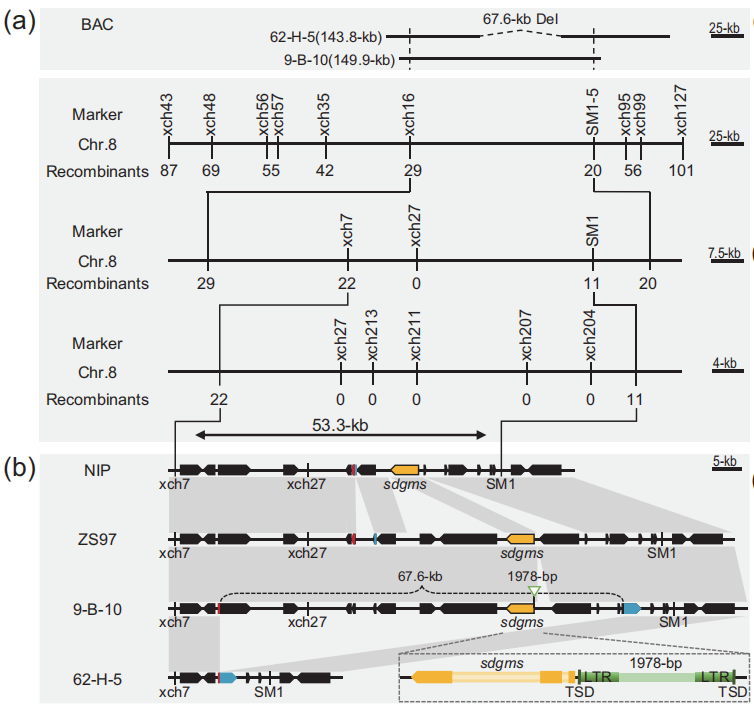
细胞核显性雄性不育材料非常少见,2001年,三明市农科院的研究者在育种材料后代中发现了一个水稻细胞核显性雄性不育的突变株,命名为SDGMS,该株系表现出稳定的完全雄性不育表型且不受环境影响,但其关键基因一直未被克隆到。
2023年8月,华中农业大学张启发院士/欧阳亦聃团队、福建省三明市农科院黄显波团队合作在National Science Review杂志上发表了题为“Spontaneous movement of a retrotransposon generated genic dominant male sterility providing a useful tool for rice breeding”的文章,首次克隆了三明显性核不育种质SDGMS中的显性雄性核不育基因SDGMS,系统地阐述了水稻核显性不育机制,为其育种应用指明了方向 (Xu et al., 2023)。
伯小远将该文的研究思路整理如下:

备注:为了方便大家理解,本文将目的基因记为sdgms,目的蛋白记为SDGMS,而SDGMS基因是指带有1978bp序列的sdgms基因,可能与原文表述有所不同,请知悉。
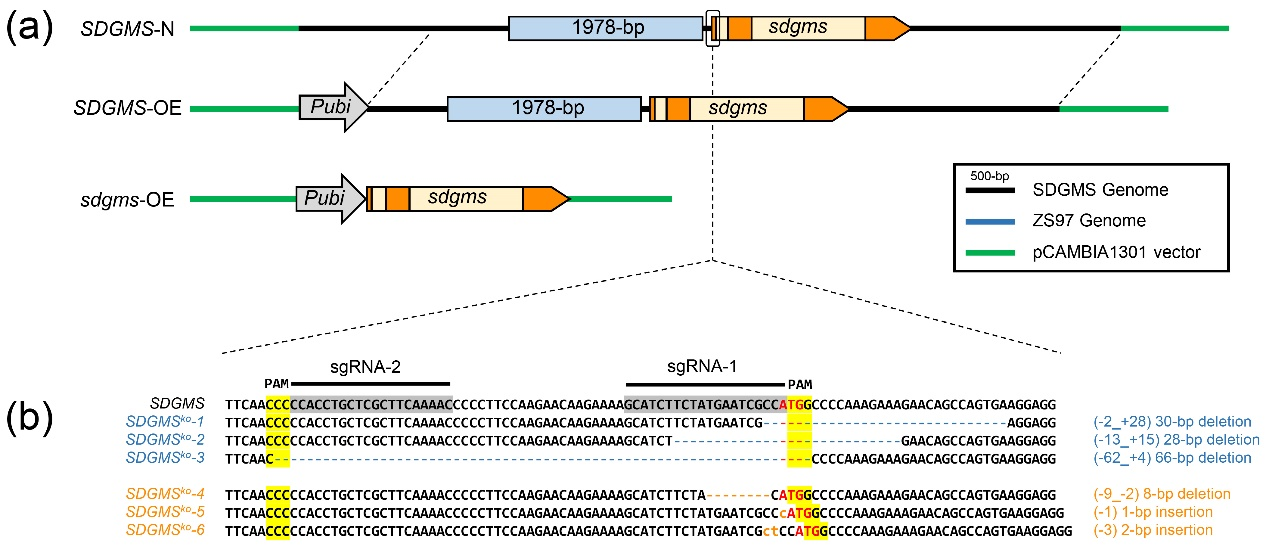
图4 载体示意图 (Xu et al., 2023)。(a)三个过表达载体SDGMS-N、SDGMS-OE和sdgms-OE;(b)敲除载体的靶点设计示意图、得到的敲除植株的具体突变形式,分为缺失起始密码子ATG的1、2、3号株系,和起始密码子ATG上游突变的4、5、6号株系。
结果显示,SDGMS-N的17个T0阳性株系表现出非常低的育性(平均13.5%);SDGMS-OE的所有26个T0阳性株系表现出完全雄性不育,无法产生花粉;sdgms-OE尽管由Ubi启动子驱动sdgms基因,同时检测到sdgms基因表达增强,但未观察到T0阳性株系育性显著降低。SDGMSko敲除株系中,缺失起始密码子ATG的3个T0株系恢复了育性(67.4%、71.7%、62.3%),而起始密码子ATG上游突变的T0株系育性未受影响(图5)。
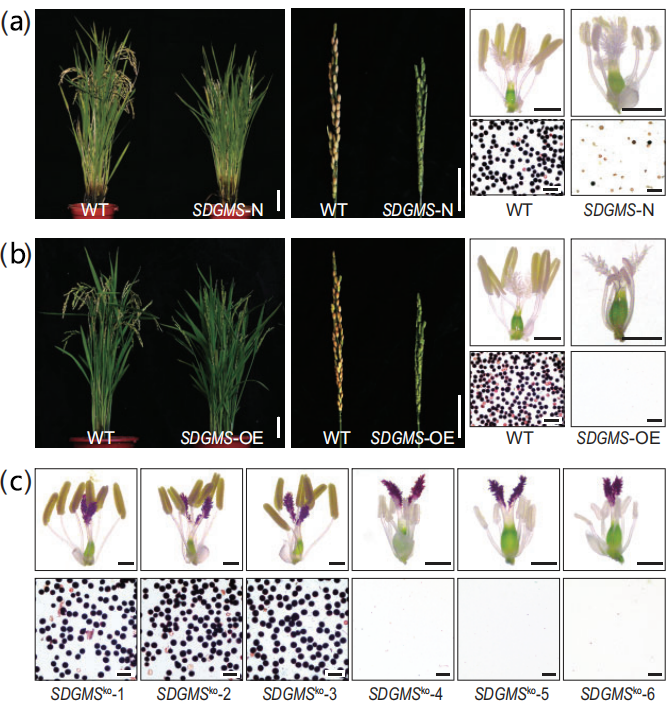
图5 对转基因植株的表型进行分析 (Xu et al., 2023)。
综上,整个互补序列对天然环境中的SDGMS品种是必需的;低于最佳长度的启动子将产生不完全的雄性不育表型,例如SDGMS-N株系;Ubi启动子可以部分补偿启动子长度的不足导致的完全不育,例如SDGMS-OE株系;没有1978bp的序列,即便是Ubi启动子也不足以使SDGMS基因在特定组织中达到不育所需的表达水平,例如sdgms基因在sdgms-OE株系中的表达量远低于其在SDGMS-OE株系中的表达量(图6)。

通过查询公共转录组数据库,发现sdgms基因在大多数品种中并无表达,仅在少数品种的幼小圆锥花序和幼穗中表达。作者团队通过qRT-PCR实验证明sdgms基因仅在不育的938(SDGMS)和ZS97(SDGMS)的幼穗第5-7期有表达(图7a)。RNA原位杂交实验检测到sdgms基因仅在ZS97(SDGMS)花药减数分裂早期的绒毡层(EM期)有表达,在ZS97(sdgms)中未被检测到(图8),与图2中异常的PCD信号一致。
作者团队使用双荧光素酶报告基因实验(LUC实验)来评估1978bp的逆转录子对基因表达的影响,发现1978bp的逆转录子产生的L/R值显著高于对照(图7b)。
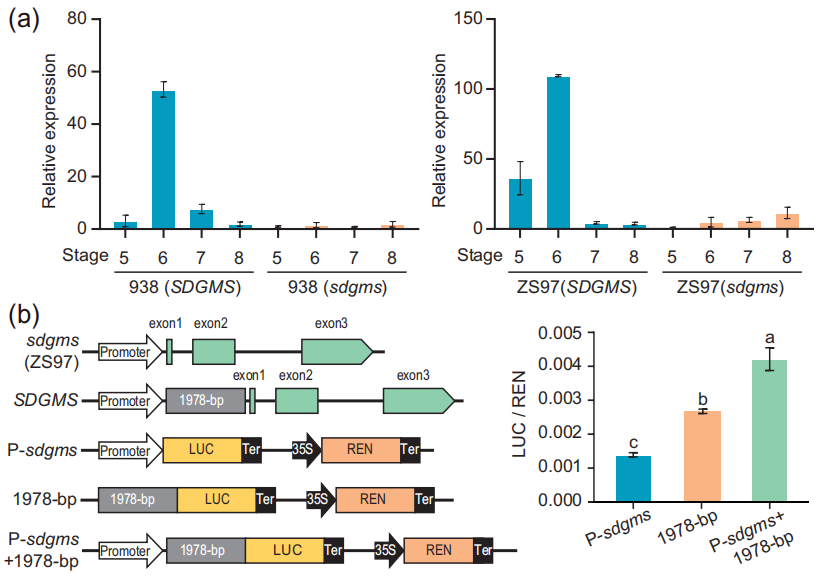
图7 sdgms基因的高表达是由1978bp的逆转录子造成的 (Xu et al., 2023)。
备注:我国学者丁颖将幼穗发育划分为8个时期:①第一苞分化期;②一级枝梗原基分化期;③二级枝梗原基及颖花原基分化期;④雌雄蕊形成期;⑤花粉母细胞形成期;⑥花粉母细胞减数分裂期;⑦花粉内容充实期;⑧花粉完成期。
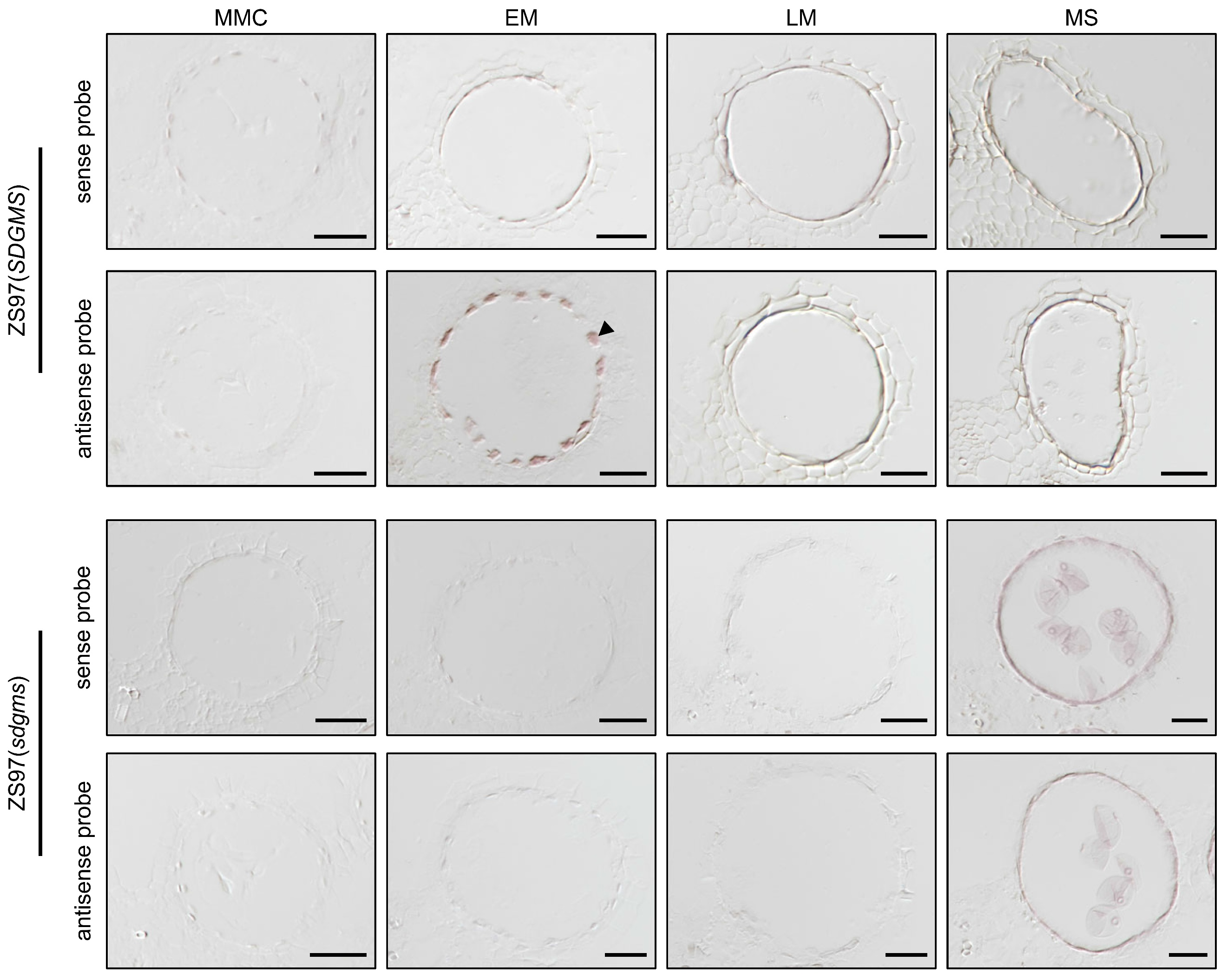
综上,在不育品种中,sdgms基因在减数分裂早期有高表达,这种高表达是由于1978bp逆转录子的插入导致的,sdgms基因的异常高表达是某些水稻品种雄性不育的主要原因。
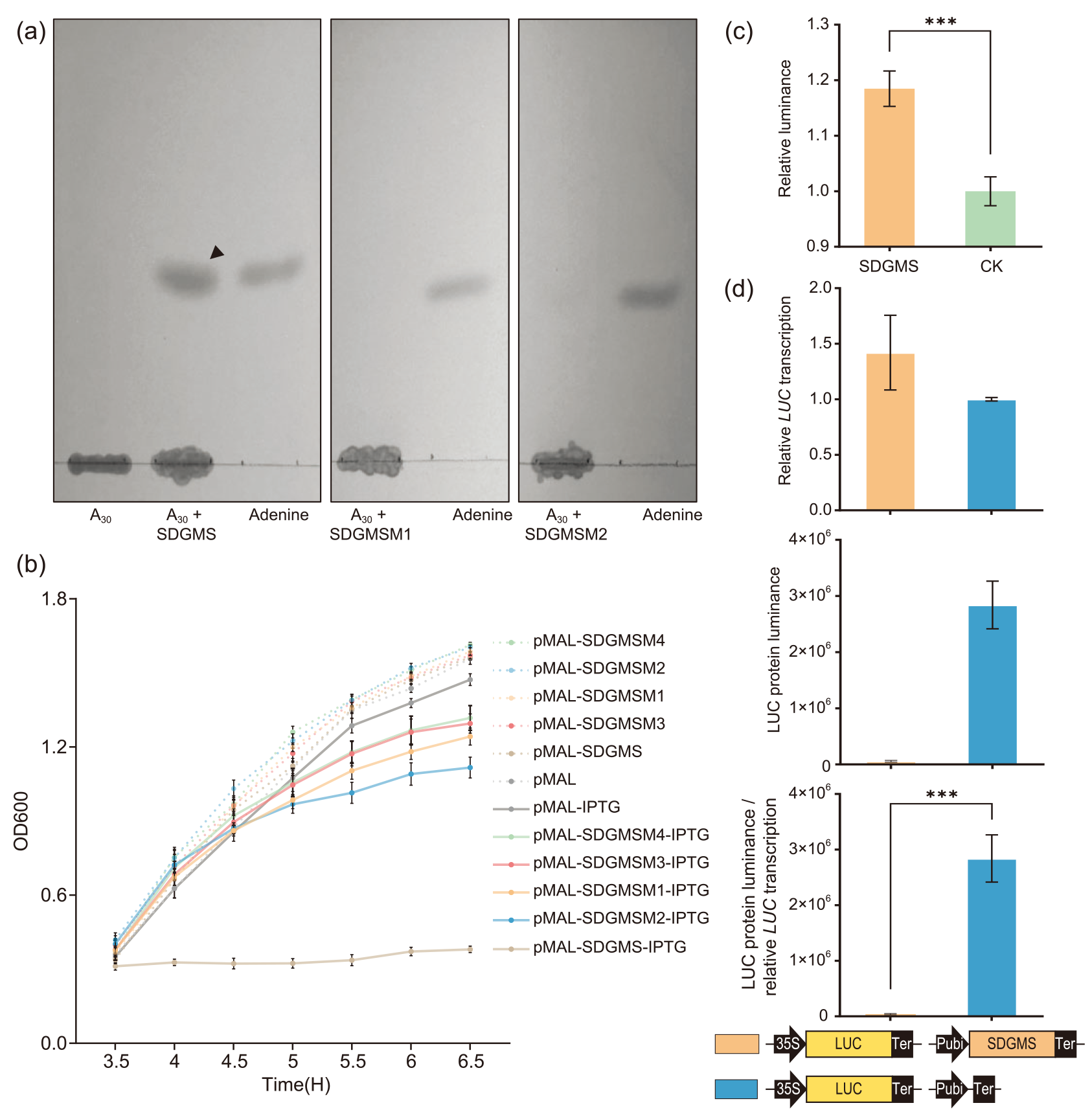
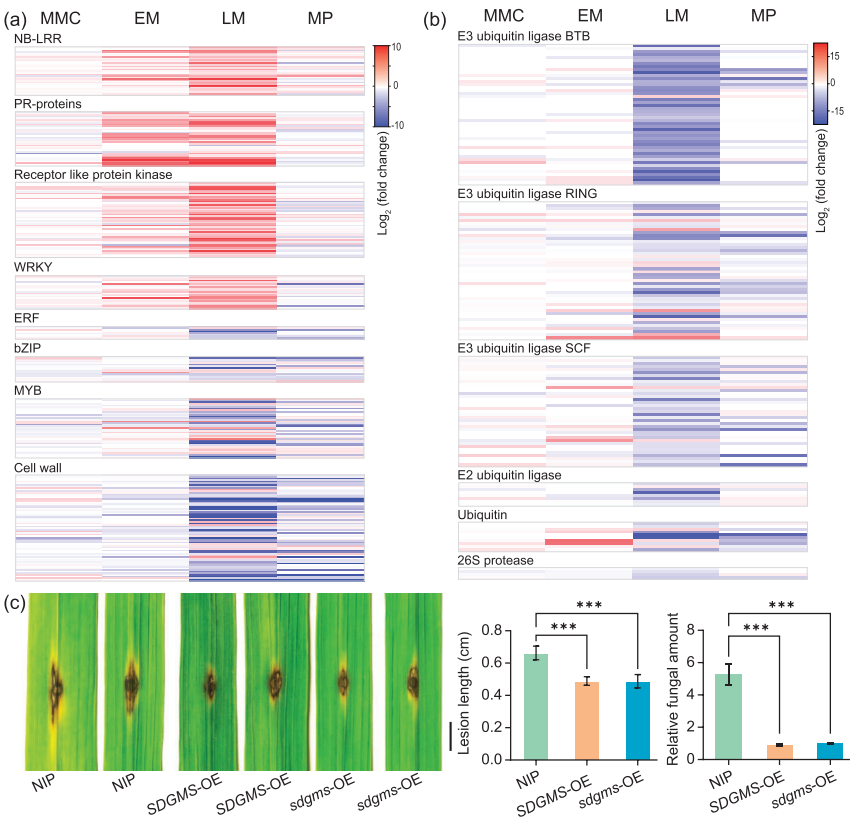
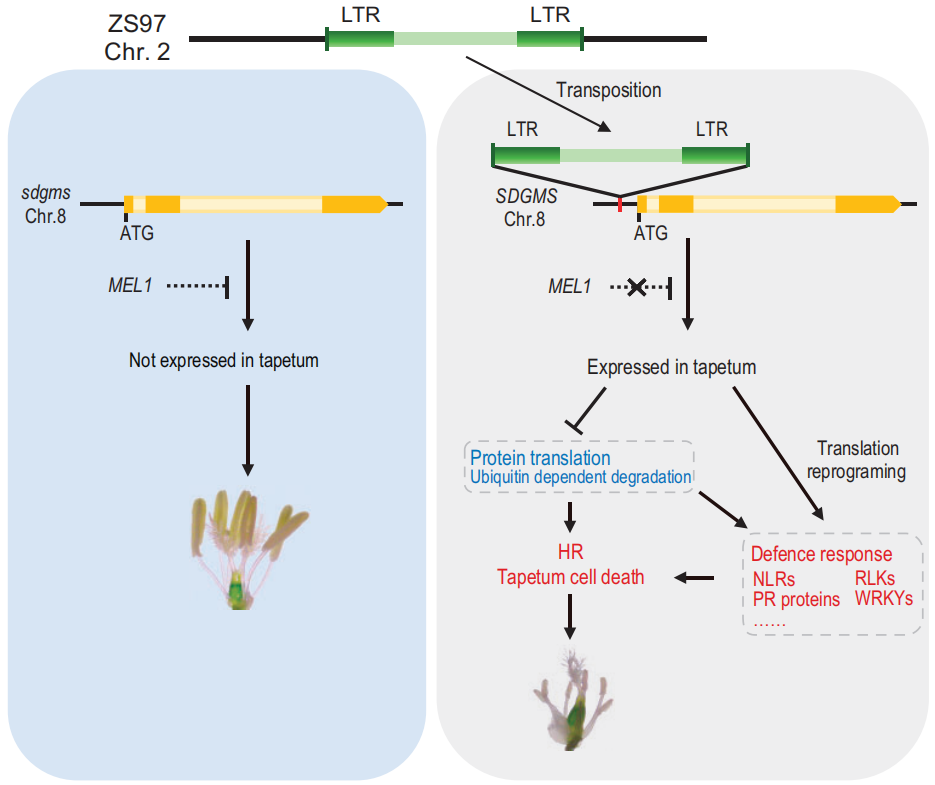
同一时间,南京农业大学、中国农业科学院万建民院士、赵志刚、朱杉杉团队在Plant Biotechnology Journal发表了题为“Anther-specific expression of OsRIP1 causes dominant male sterility in rice”的文章,首次定位出三明-Dms水稻(Dms1水稻)细胞核显性雄性不育株系中的关键作用基因OsRIP1,并对其导致不育的分子机理进行了解析,为今后利用显性不育材料进行育种指明了方向 (Lei et al., 2023)。
伯小远将该文的思路整理如下:

图13 与NJ4相比,NIL-NJ4Dms1的花药要小得多 (Lei et al., 2023)。
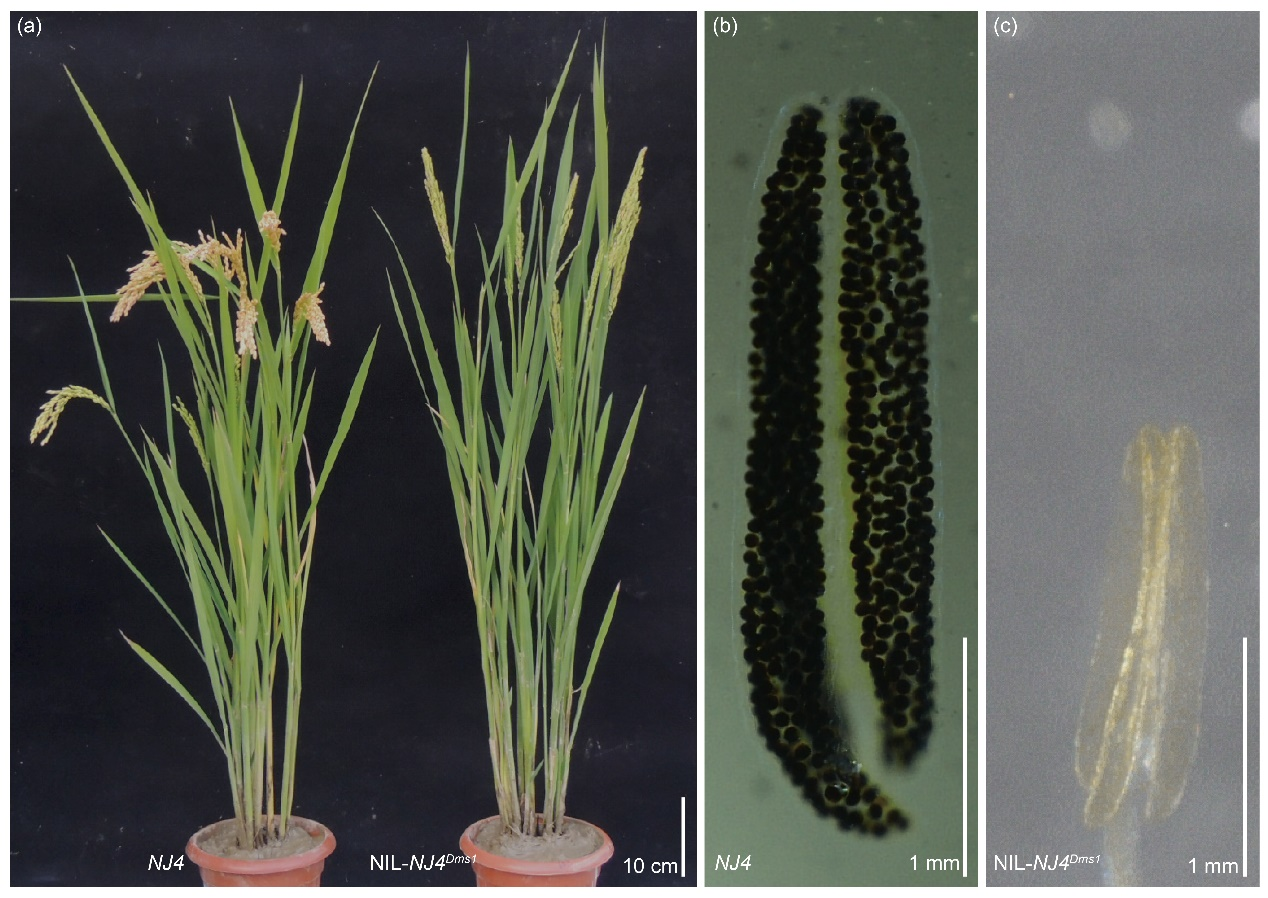
图14 NJ4与NIL-NJ4Dms1之间在形态学上无明显差异,碘-碘化钾染色试验显示NIL-NJ4Dms1中无花粉 (Lei et al., 2023)。
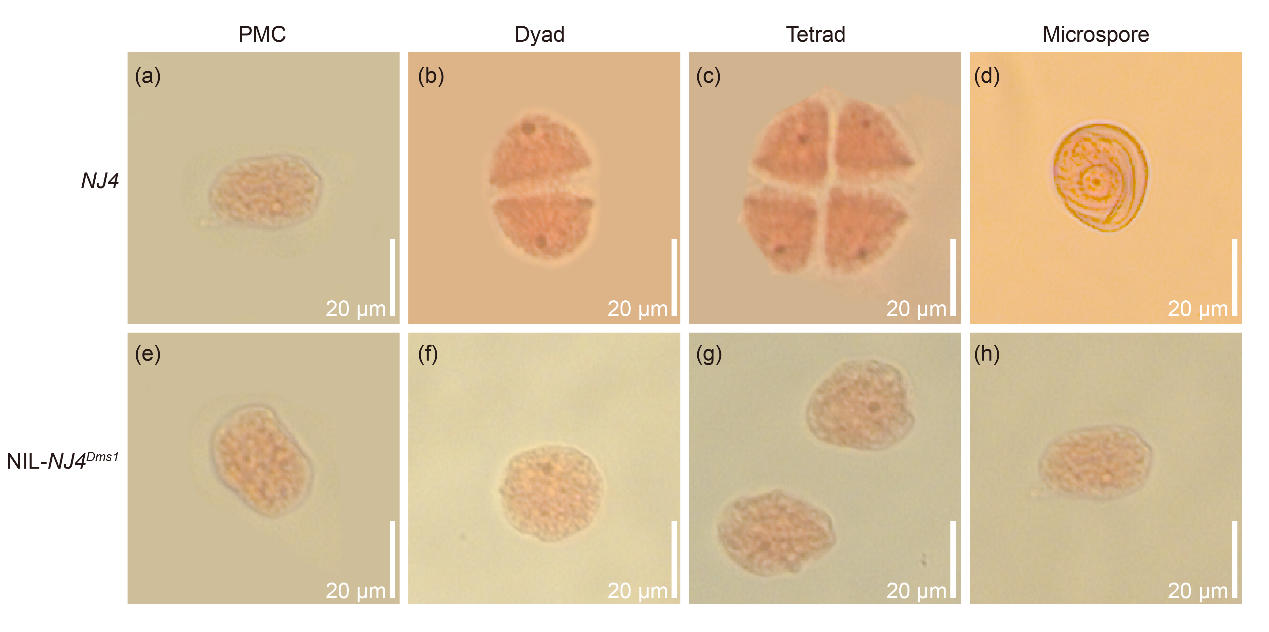
图15 胭脂红染色显示NIL-NJ4Dms1中花粉母细胞减数分裂存在缺陷 (Lei et al., 2023)。
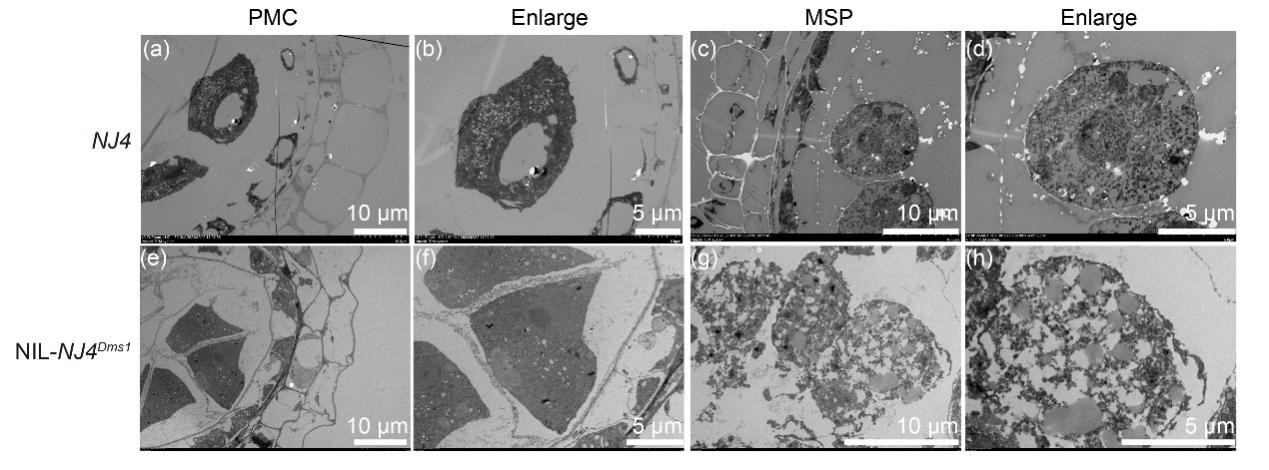

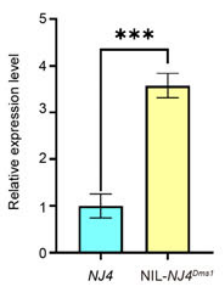
图18 qRT-PCR检测OsRIP1在NJ4和NIL-NJ4Dms1中的表达情况 (Lei et al., 2023)。
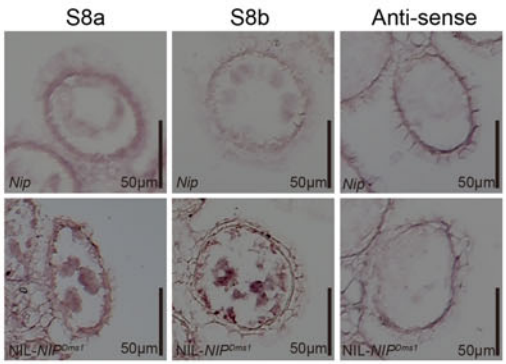
分别克隆NJ4和NIL-NJ4Dms1中OsRIP1的启动子,发现在NIL-NJ4Dms1中OsRIP1的起始密码子上游98bp处有一个1978bp的片段插入。分别构建启动子::GUS载体,转化后检测OsRIP1的时空表达特性,结果显示,GUS在pRIP1Dms1::GUS植株中均有表达,而在pRIP1Nip::GUS植株中没有表达(图19、图20)。
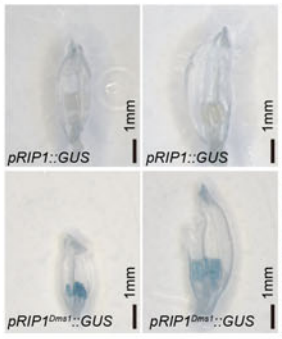
图19 检测OsRIP1在小穗中的表达 (Lei et al., 2023)。

图20 检测OsRIP1的表达 (Lei et al., 2023)。
原位杂交检测OsRIP1在花药的表达情况,结果与上述实验相同(图21)。
使用花药特异性启动子EAT1驱动OsRIP1基因,转入日本晴后,出现了类似NIL-NJ4Dms1的不育性状(图22b)。分别使用特异性根、特异性叶片启动子驱动OsRIP1基因,转入日本晴后,植株会出现死亡(见原文补充材料)。
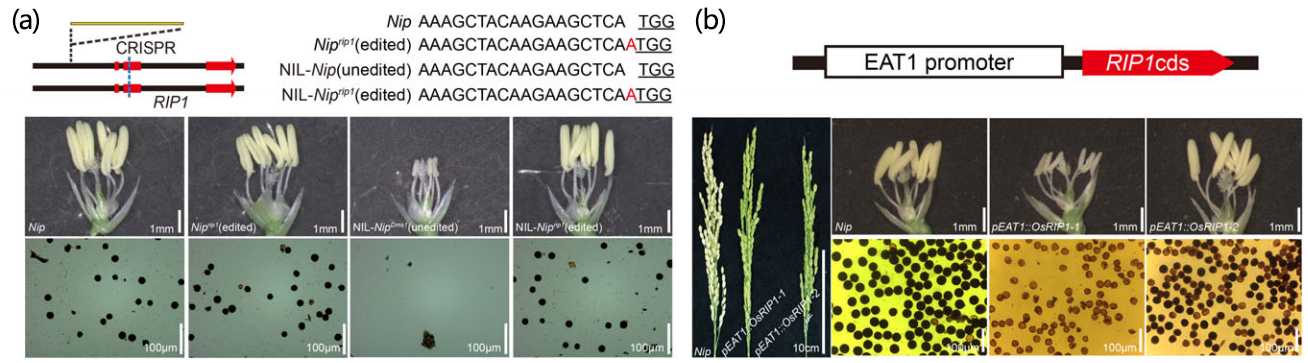
以上结果表明,OsRIP1的强表达是导致Dms株系不育的原因。

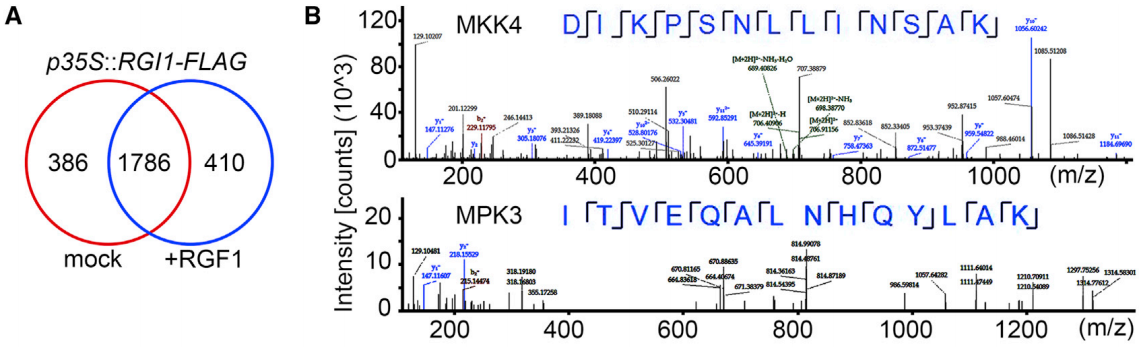
作者团队进行了IP-MS实验,GO分析表明OsRIP1可能通过抑制核糖体的翻译功能从而产生毒性(图23e)。体外翻译抑制实验也证明了这个猜想(图23f、g)。
以上实验表明,OsRIP1通过抑制核糖体的翻译功能而不是通过去嘌呤化来毒害植物细胞。
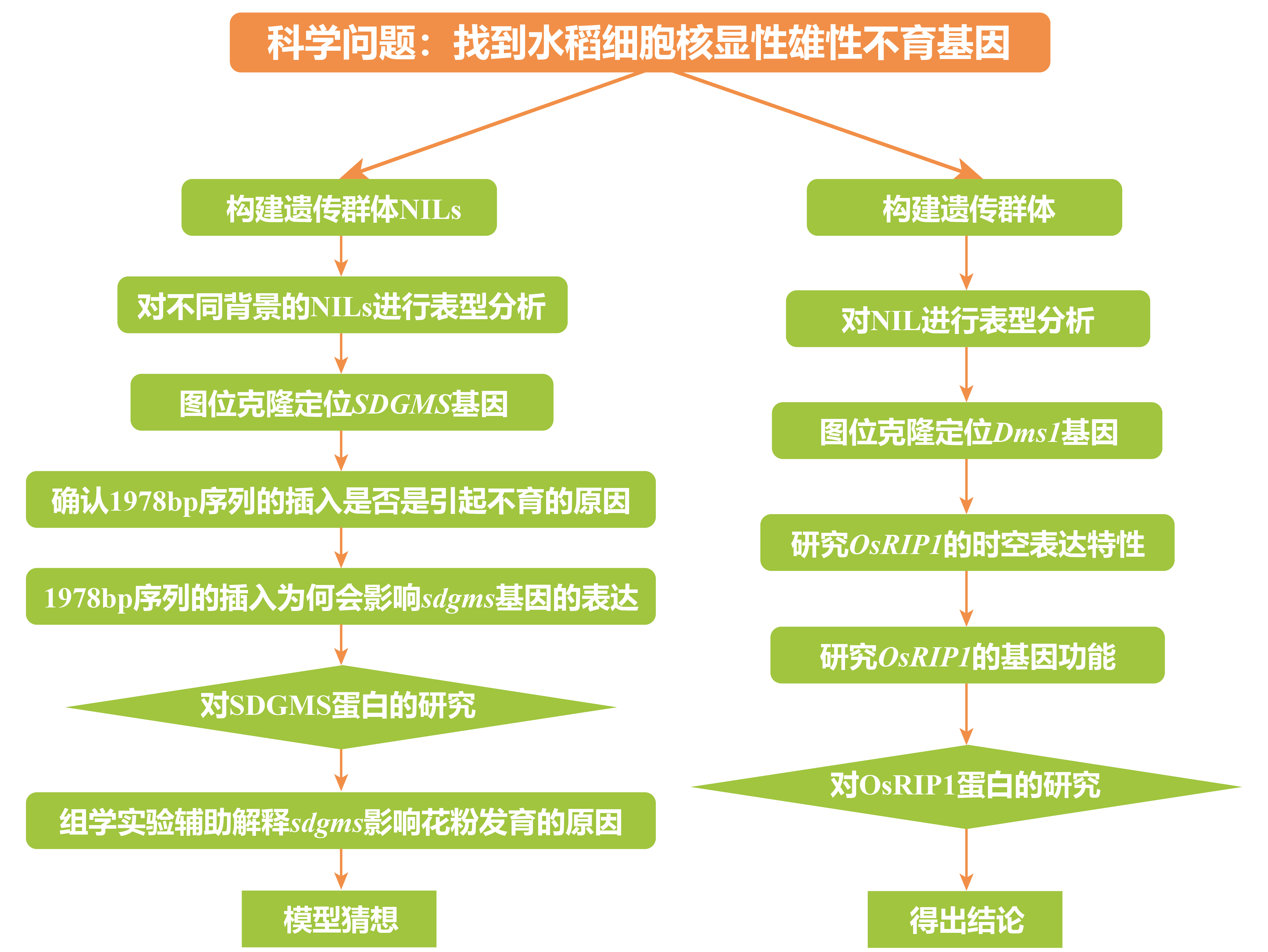
我们仔细阅读两篇论文,可以发现它们整体的研究思路非常一致,只是许多实验细节有很大不同,比如,对不育材料表型的分析、对目的基因功能的分析、对目的蛋白功能的分析,两个团队都各自使用了不同的实验方法。
再说说各自的优点。两个团队都发现了一个1978bp的序列在目的基因启动子上的插入,但第一个团队对这个序列进行了更为详细的分析,根据其结构判断其为反转录转座子,而且发现了其来源于亲本ZS97,也使用了定量的方法来分析该序列对sdgms基因表达的影响。并且该团队对sdgms基因最终导致不育的原因也试图进行了更为详细的解释,即sdgms基因可能触发了花粉内源性的应激反应导致绒毡层PCD异常。
第二个团队实验设计非常明朗、清爽、引人入胜,实验均是围绕着NJ4和NIL-NJ4Dms1两种材料来进行,而且通篇基因仅记为Dms1/OsRIP1,对读者阅读并理解论文的内容十分友好,是让人在阅读中停不下来的“爽文”。
不容忽略的是,两篇论文中有一个结论是完全相反的(图9和图23),即SDGMS/RIP1这个核糖体失活蛋白是否具有去嘌呤化的作用,伯小远猜测是由于实验测试系统的不同导致得出了不同的结论,一个团队使用的是原核表达系统,另一个团队使用的是真核表达系统。大家如果感兴趣的话,还是要去读读两篇原文喔!
2020年9月,兰州大学黎家团队在Molecular Plant杂志上发表了题为“RGF1-RGI1, a peptide-receptor complex, regulates Arabidopsis root meristem development via a MAPK signaling cascade”的文章,作者团队通过遗传学、生化实验及细胞生物学实验最终证明,RGF1-RGI1-YDA-MKK4/MKK5-MPK3/MPK6-PLT1/PLT2级联信号通路调控拟南芥根分生组织发育 (Lu et al., 2020)。
伯小远将该文的思路整理如下:



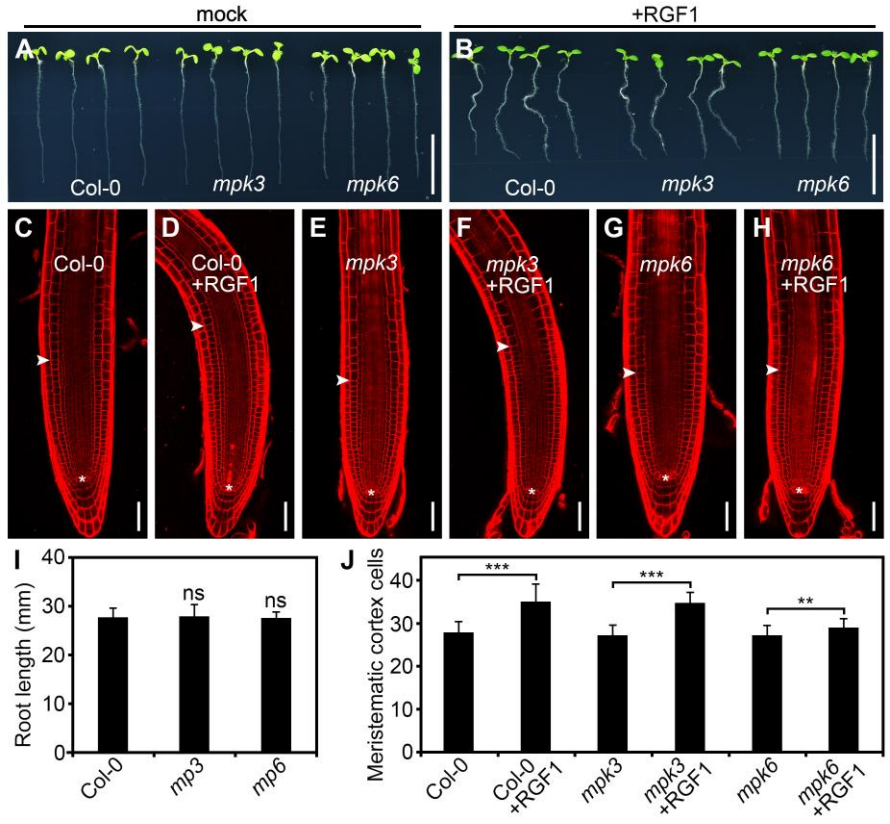

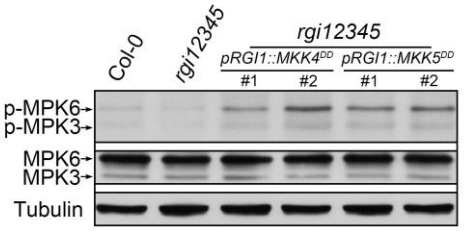
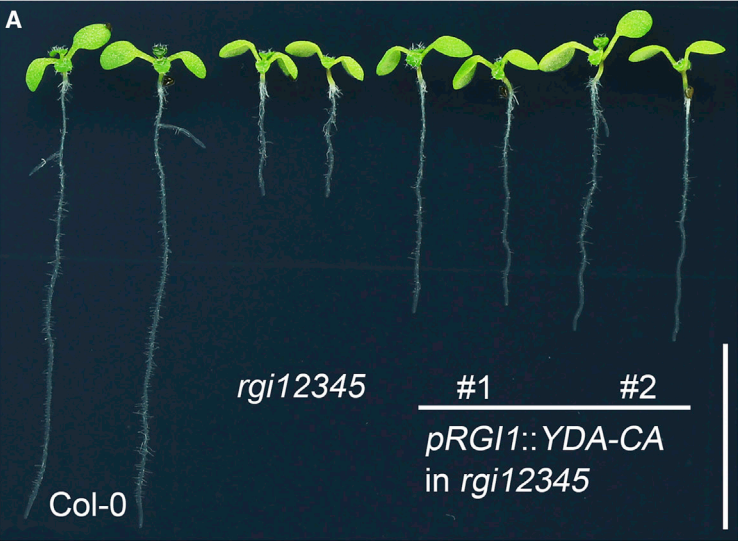
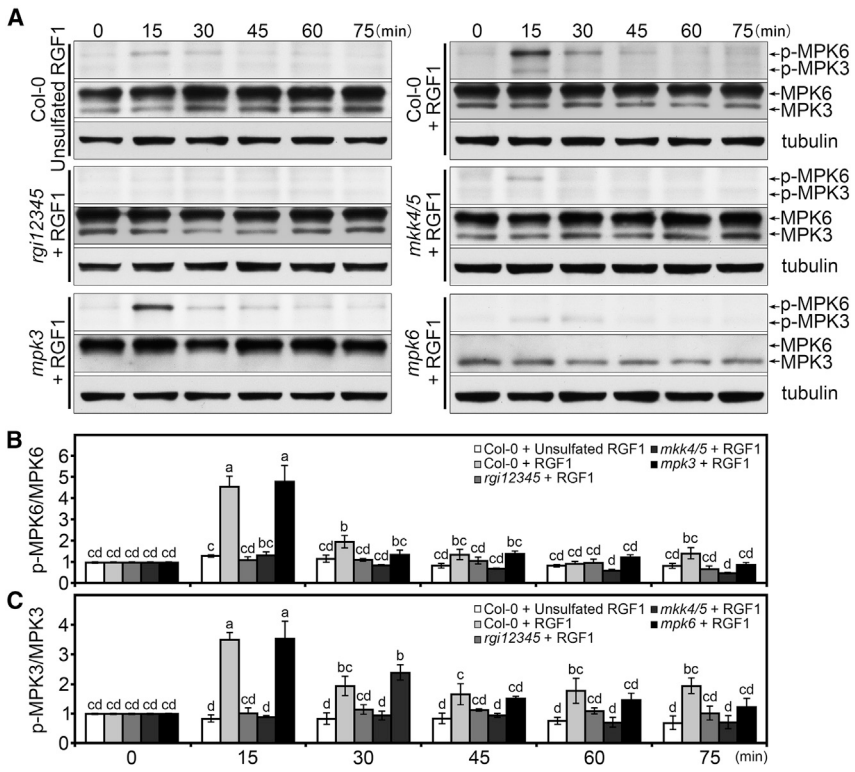

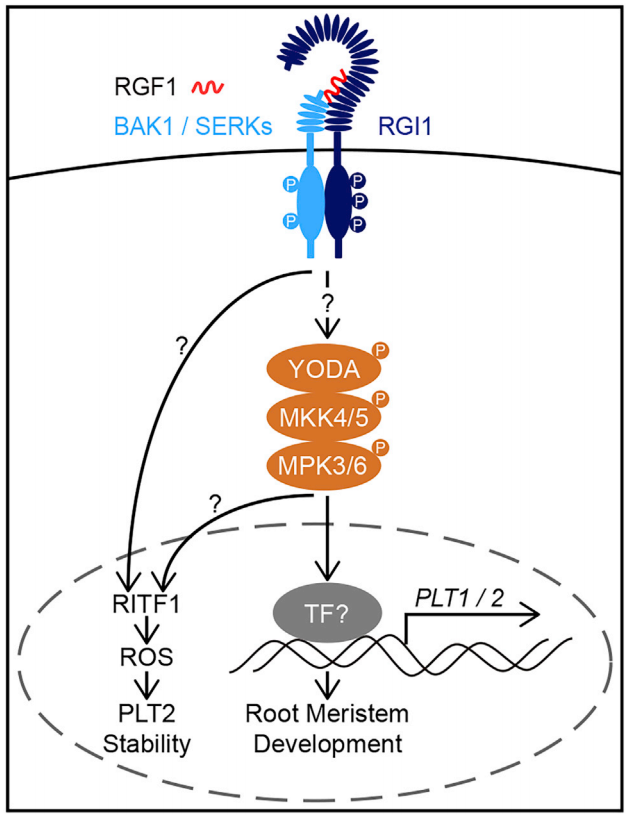
同一时间,浙江大学徐娟团队与美国密苏里大学张舒群团队合作,和上篇文章在Molecular Plant杂志上以背靠背方式发表了题为“The YDA-MKK4/MKK5-MPK3/MPK6 cascade functions downstream of the RGF1-RGI ligand-receptor pair in regulating mitotic activity in the root apical meristem”的文章,该团队长期致力于研究MAPK级联信号在植物生长发育中的功能,最终也证明了RGF1-RGI1-YDA-MKK4/MKK5-MPK3/MPK6-PLT1/PLT2级联信号通路在调控拟南芥根分生组织发育中的重要作用 (Shao et al., 2020)。
篇幅所限,伯小远将这篇论文的研究思路总结如下:
从三种与RAM有关的转录因子SHR-SCR、WOX5和PLT1/PLT2中筛选出PLT1/PLT2位于MPK3和MPK6下游。
由于MAPKK蛋白如MKK4、MKK5、MKK7和MKK9被报道过是MPK3/MPK6上游蛋白,因而鉴定各mkk突变体在RAM发育中的作用,结果显示MKK4和MKK5与RAM发育相关。
以上实验发现MKK4/MKK5-MPK3/MPK6信号通路功能缺失会导致植物主根变短,与RGF1-RGI信号通路缺失表型类似,因此着手研究MKK4/MKK5-MPK3/MPK6信号通路与RGF1-RGI信号的上下游关系,检测RGF1对MPK3/MPK6磷酸化的影响,结果显示RGF1诱导的MPK3/MPK6磷酸化依赖于RGI受体和MKK4/MKK5。
为进一步研究MKK4/MKK5-MPK3/MPK6信号通路与RGF1-RGI信号的关系,将RGI2基因启动子驱动持续激活的MKK4/MKK5转入rgi12345突变体。
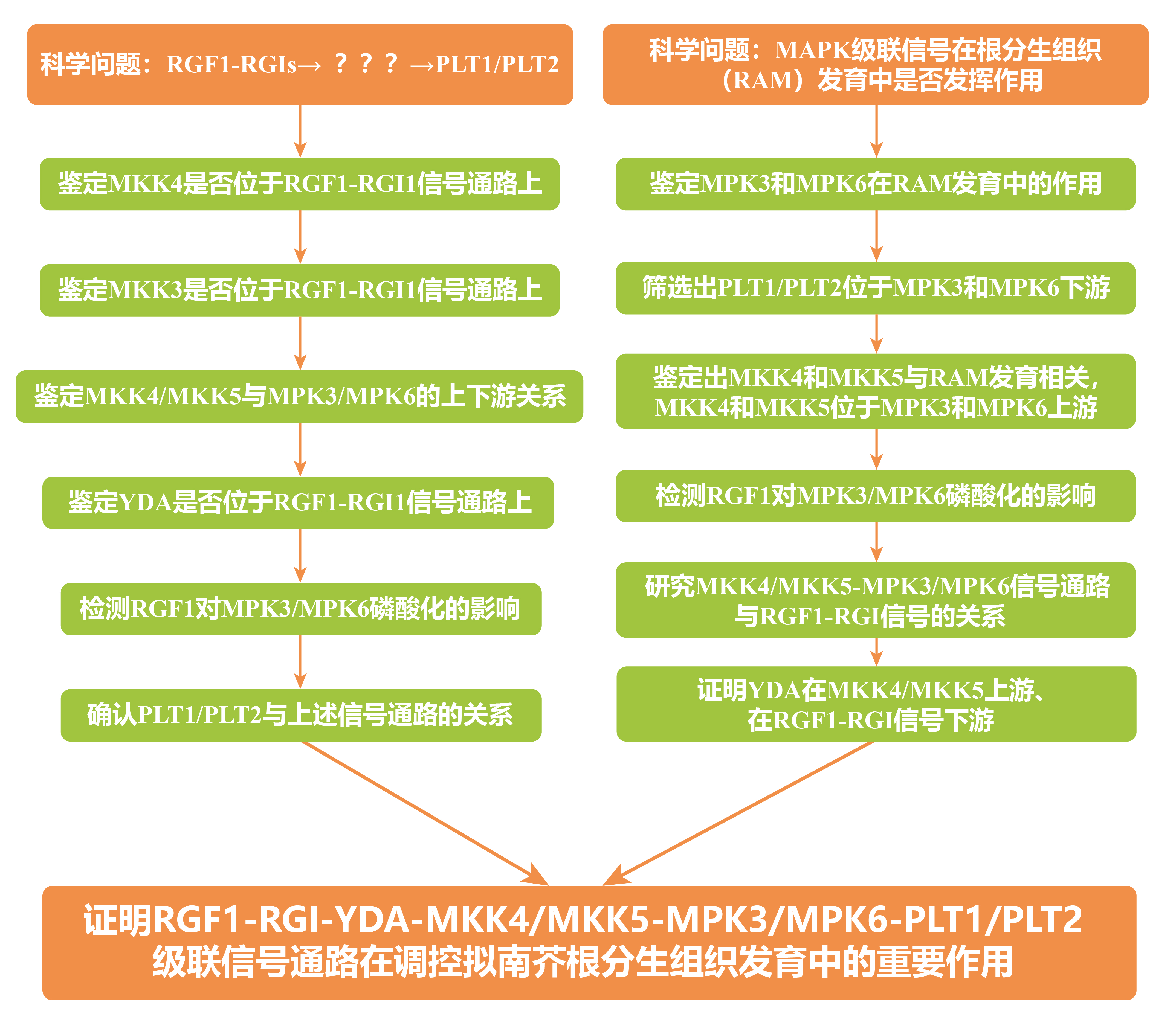
例子二中的这两篇研究论文从不同角度出发,最终得出了共同的结论。根分生组织发育涉及到非常精细复杂的调控网络,不仅包括信号通路上基因的鉴定,也包括了各基因在信号通路中的上下游关系。第一个团队已知级联信号的两端,为了探索中间的信号通路,以蛋白组学技术IP-MS为突破口,先找到相互作用蛋白,再逐级进行验证。第二个团队由级联信号的末端,逐级向上不断寻找并串联起了整个级联信号途径。两个团队的分子实验都做的非常深入,尤其是利用遗传学实验、生化实验和细胞生物学实验证明两个基因的上下游关系,设计得都非常严谨,很值得大家学习。
伯小远在本文列举了两个发表背靠背论文的例子,第一个例子是为了解决相同的科学问题设计了不同的实验并得出了相同的结论,第二个例子是为了解决不同的科学问题但最终证明了相同的级联信号通路。背靠背论文是我们学习不同的科研思考角度不可多得的好资料,感谢这些作者团队为我们读者献上了不可多得的科研盛宴!更多内容等你来探索!
References:
Lei D, Jian A, Huang X, et al. Anther-specific expression of OsRIP1 causes dominant male sterility in rice. Plant Biotechnol J. 2023;21(10):1932-1934. doi:10.1111/pbi.14140
Lu X, Shi H, Ou Y, et al. RGF1-RGI1, a Peptide-Receptor Complex, Regulates Arabidopsis Root Meristem Development via a MAPK Signaling Cascade. Mol Plant. 2020;13(11):1594-1607. doi:10.1016/j.molp.2020.09.005
Shao Y, Yu X, Xu X, et al. The YDA-MKK4/MKK5-MPK3/MPK6 Cascade Functions Downstream of the RGF1-RGI Ligand-Receptor Pair in Regulating Mitotic Activity in Root Apical Meristem. Mol Plant. 2020;13(11):1608-1623. doi:10.1016/j.molp.2020.09.004
Xu C, Xu Y, Wang Z, et al. Spontaneous movement of a retrotransposon generated genic dominant male sterility providing a useful tool for rice breeding. Natl Sci Rev. 2023;10(9):nwad210. Published 2023 Aug 7. doi:10.1093/nsr/nwad210
