单碱基突变可以引起许多农艺性状的改变,最常见的原因是基因编码区内单碱基突变导致氨基酸发生突变,进而导致蛋白的功能发生改变,从而产生新的性状。现在常通过基于CRISPR-Cas9/Cas12a的单碱基编辑器ABE、CBE或PE系统实现对目标基因的单碱基突变,用以人工创制新种质资源。今天伯小远打算写一写在自然变异或通过EMS创制的突变体中,单碱基突变有哪些类型,是不是不止刚才说的编码区内的单碱基突变这一种情况呢,下面我们来一起看看吧。
本次推文的主要内容是:
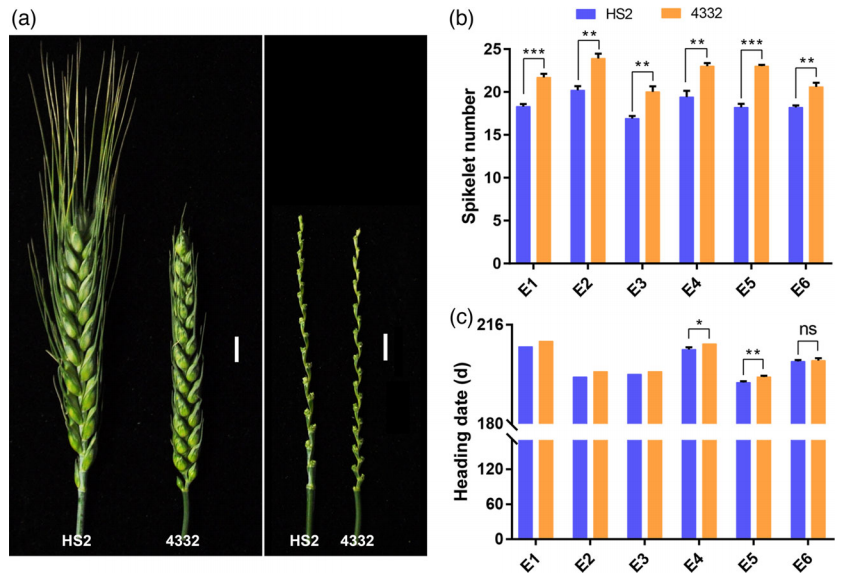
图1 HS2和4332的穗形态(a)、小穗数(b)、抽穗期(c)的统计 (Chen et al., 2022)。
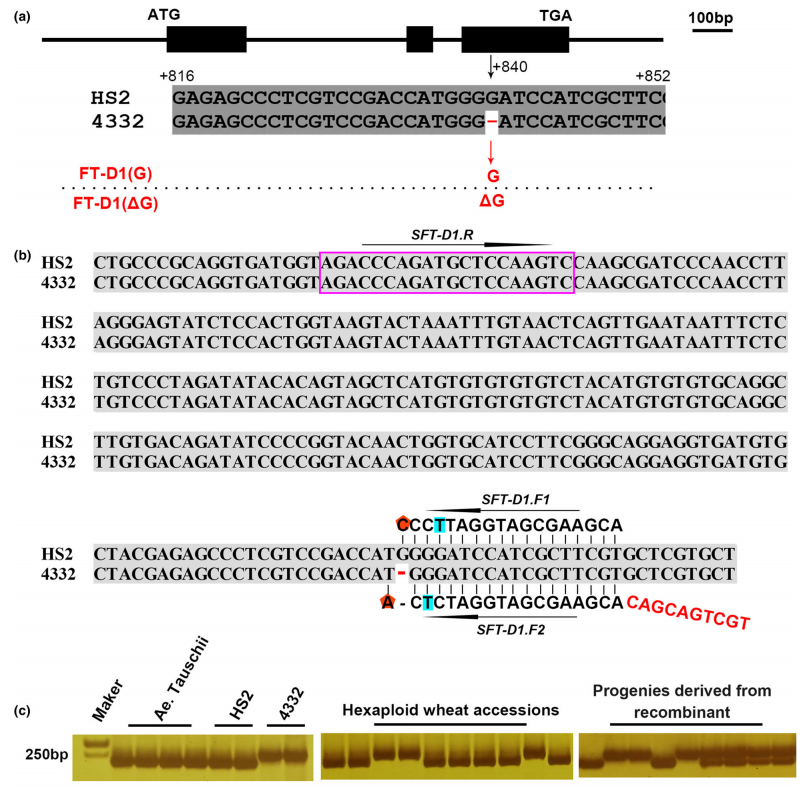
图2 FT-D1基因的多态性及STARP标记开发 (Chen et al., 2022)。作者通过小穗数和抽穗期差异显著的材料HS2和4332定位出与这两个性状有关的QTL在染色体7DS一段60M的区间内,再通过精细定位缩小至2.7Mb的区间内,在该区间内包含了一个著名的开花基因FT,通过对HS2和4332材料中该基因的测序发现FT在两个材料中存在一个碱基G的差异,位于第三个外显子中(a),将其命名为FT-D1,通过分子标记证明FT-D1是目标基因。根据FT-D1基因的多态性设计了STARP标记SFT-D1(b),根据该标记扩增不同小麦品种可以快速、经济地预测小穗数和抽穗期(c)。
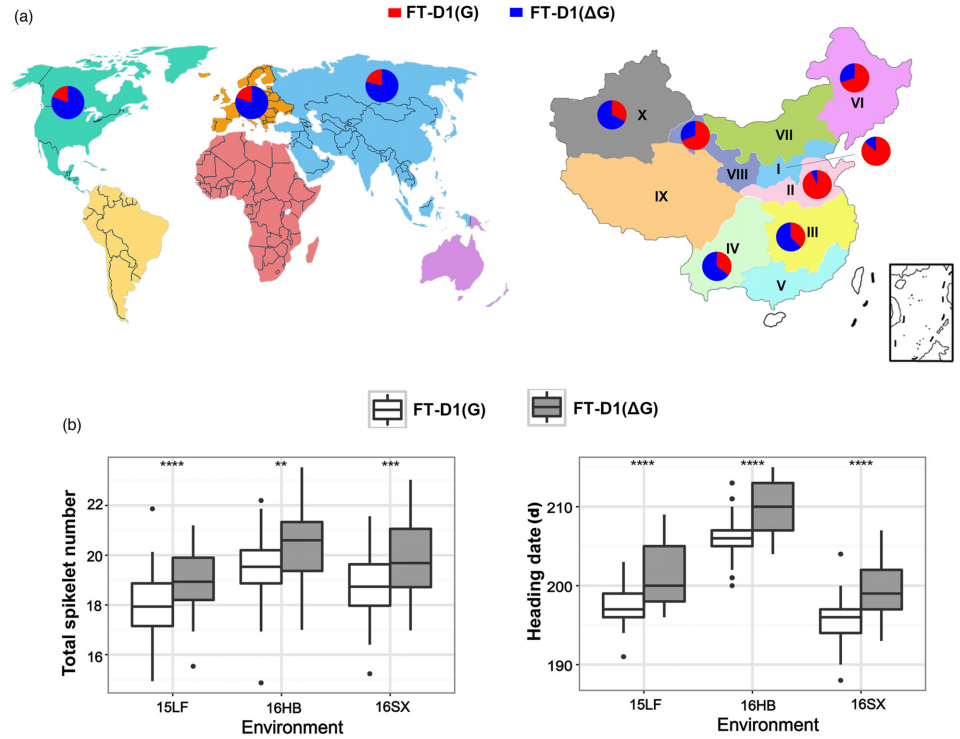
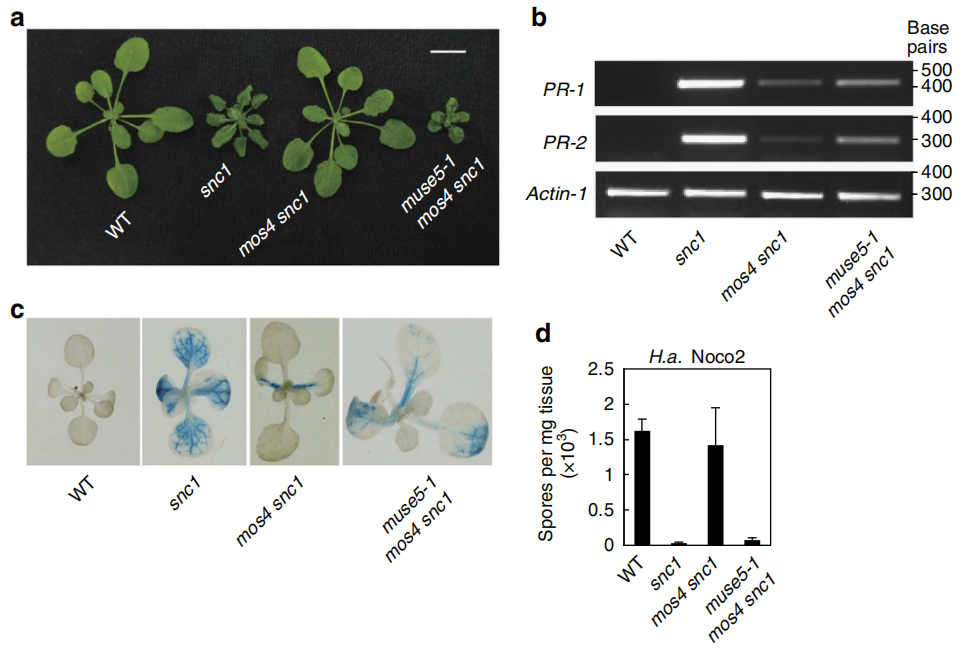

图4 过表达SlWRKY33或ShWRKY33 (Guo et al., 2022)。该团队通过转录组数据发现对冷敏感的栽培番茄S. lycopersicum L ‘Ailsa Craig’中SlWRKY33基因不会被低温诱导表达,而耐寒的野生番茄S. habrochaites LA1777、S. pennellii LA0716中的ShWRKY33、SpWRKY33基因会被低温强烈诱导表达。分析SlWRKY33和ShWRKY33蛋白发现有6个氨基酸的差异,因而将‘Ailsa Craig’作为受体材料,转入35S:SlWRKY33(图中SlW33-OE#1/2材料)或35S:ShWRKY33(图中ShW33-OE#1/2材料),结果显示,这些过表达材料都增强了番茄的耐寒性,从这个结果可推测造成栽培番茄和野生番茄耐寒性的不同是由于WRKY33的表达模式的差异而不是其蛋白功能的差异。

图5 对SlWRKY33和ShWRKY33的启动子区域进行转录活性分析 (Guo et al., 2022)。作者通过双荧光素酶报告基因实验(LUC实验)发现SlWRKY33和ShWRKY33的启动子中有一个关键顺式作用元件W-box序列发生了一个单碱基变异(a、b),前者记为pSlW33,序列为TTGATT,后者记为pShW33,序列为TTGACT。冷胁迫下,通过LUC实验发现将pSlW33序列突变为mpSlW33后LUC/REN的值显著升高,而将pShW33序列突变为mpShW33后LUC/REN的值与对照无差异(c、d),更加确认了该W-box序列中发生的单碱基变异造成了SlWRKY33和ShWRKY33转录的差异。EMSA、ChIP-qPCR实验证明W-box序列pShW33可以与SlWRKY33或ShWRKY33蛋白结合,但W-box序列pSlW33却不能。根据以上实验结果推测,在冷胁迫下,野生番茄中的ShWRKY33蛋白可自激活ShWRKY33基因启动子中的W-box,但栽培番茄中由于SlWRKY33基因启动子中的W-box发生了单碱基变异,SlWRKY33无法结合自身基因SlWRKY33启动子。
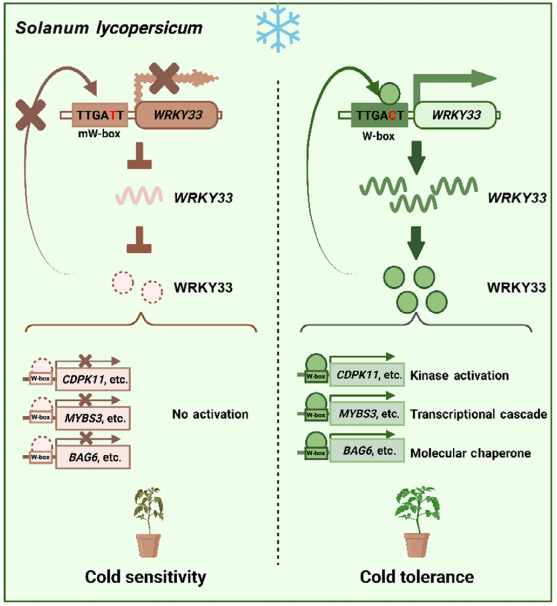

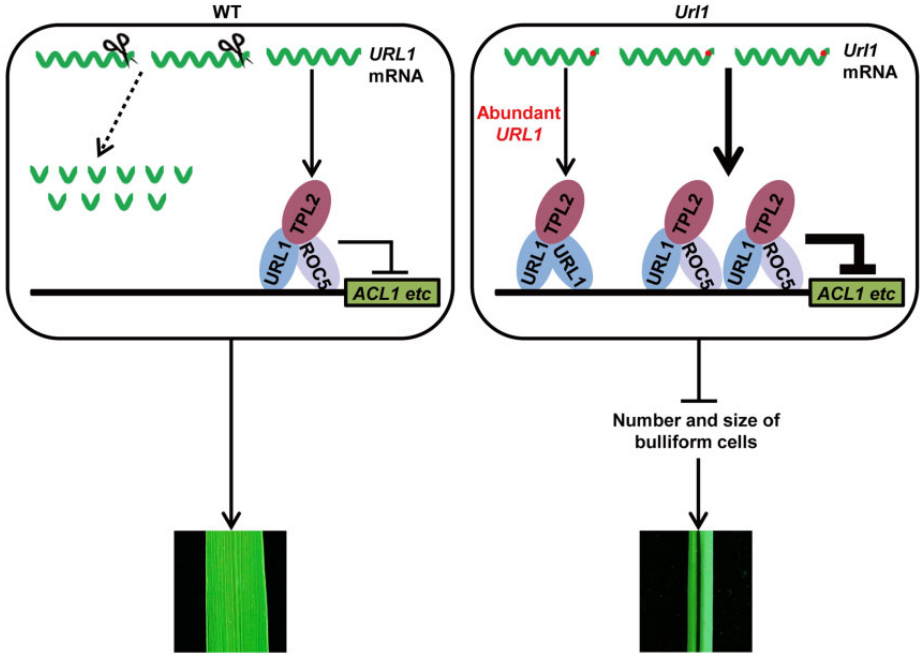
图7 Url1突变体表型分析 (Fang et al., 2021)。作者发现与WT的扁平叶片相比,Url1突变体的叶片卷成圆筒状,F1的叶片呈半卷状(A-C),用甲苯胺蓝O染色发现在WT中,沿大叶脉和小叶脉被染成紫色的泡状细胞排列成三列,在Url1突变体中只观察到两列(D-G),WT和Url1成熟叶片的横切面(H-M),WT和Url1的LRI(N)、泡状细胞数量(O)和面积(P)。备注:泡状细胞数量和大小的减少会使叶片卷曲。
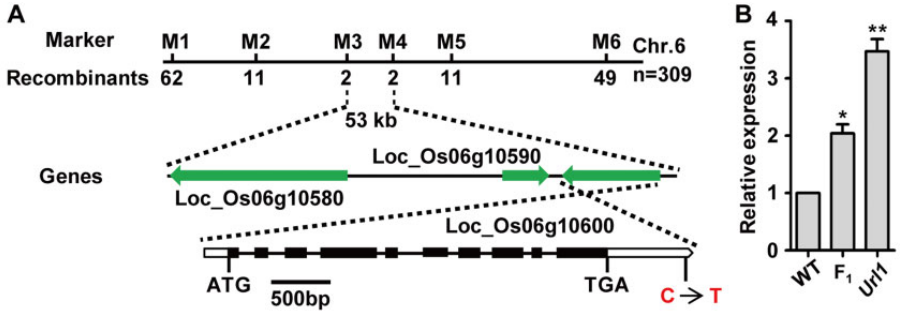
图8 URL1基因的图位克隆 (Fang et al., 2021)。作者利用图位克隆技术鉴定出在突变体Url1中LOC_Os06g10600的终止密码子后第679nt处发生了一个单碱基替换(A),该突变位点位于LOC_Os06g10600的3’-UTR区,将LOC_Os06g10600命名为URL1,该基因编码HD-ZIP IV家族转录因子ROC8。在Url1突变体中,URL1的3’-UTR中保守基序的单碱基突变,增强了URL1 mRNA的稳定性,使得URL1基因在突变体中的表达量显著高于野生型(B)。
2013年,哥伦比亚大学Li Xin团队在Nature communications杂志上发表了题为“Mitochondrial AtPAM16 is required for plant survival and the negative regulation of plant immunity”的研究论文,该团队通过EMS化学诱变筛选出拟南芥snc1增强子突变体muse5,并且发现MUSE5和酵母PAM16同源,定位于线粒体内膜,首次揭示了拟南芥MUSE5/TXR1/AtPAM16负向调控R蛋白SNC1介导的免疫反应,再次证明了线粒体参与植物免疫反应,拓展了对植物R蛋白介导的免疫信号的认识 (Huang et al., 2013)。

图10 muse5-1的图位克隆 (Huang et al., 2013)。拟南芥SNC1是一种R蛋白,snc1突变体使SNC1异常活跃,可抗病原菌,植株矮小。mos突变体为snc1抑制子突变体,这些突变体在表型上与野生型基本一致,但也抑制了snc1的抗病性。作者想筛选出snc1增强子突变体,因此通过EMS诱变mos snc1双突突变体,得到了类似于snc1突变体的muse突变体,植株矮小且抗病。作者通过群体进行图位克隆,最终克隆到At3g59280发生了突变,突变的位置为内含子与外显子的剪切处,碱基G突变为A(a、b),对突变体的cDNA与野生型比较,发现碱基G在muse5-1中缺失(c),最终导致MUSE5蛋白改变。通过比对蛋白序列,发现MUSE5与酿酒酵母中的PAM16有35%的相似度,均存在J-Like保守结构域(e虚线处)。
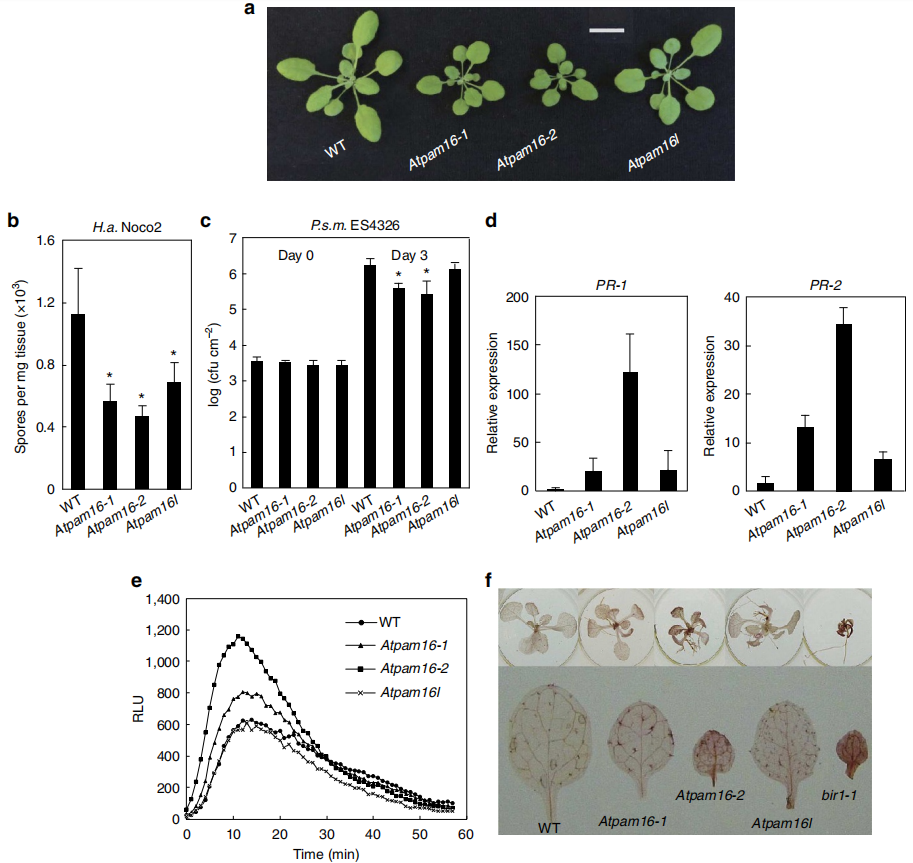
图11 AtPam16突变体的抗病性分析 (Huang et al., 2013)。基因AtPAM16(即MUSE5)突变,造成mos4 snc1抗病性恢复,所以推测AtPAM16可能是一个免疫负调控因子,从表型上看,AtPam16-1 (muse5-1)与AtPam16-2(txr1-1,txr1-1是前人报道的At3g59280基因的突变体)均小于野生型,AtPam16l(At5g61880 T-DNA突变体,At5g61880编码产物与MUSE5类似)与野生型类似(a);从抗病性检测来看,结果可见图b、c。综合来看,基因AtPAM16突变引起了拟南芥的抗病性,因此AtPAM16是一个免疫负调控因子。

图13 mdm-siR277-1/2通过调控MdRNL1/2/3/4/5基因的表达影响苹果的ALT1易感性 (Zhang et al., 2018)。作者在感病品种GD叶片中接种ALT1,发现mdm-siR277-1/2被诱导(A),接种2h后其靶基因MdRNL1/2/3/4/5的mRNA水平相应降低(B),接着利用STTM沉默mdm-siR277-1/2(C、D),发现其靶基因MdRNL1/2/3/4/5表达量增加(E),并且发现叶片感染症状减轻(F)、感病率降低(G)。随后作者在抗病品种HF叶片中接种ALT1,发现mdm-siR277-1/2未被诱导(H),并且其靶基因MdRNL1/2/3/4/5的表达升高(I),接着在抗病品种HF中超表达MdhpRNA277后发现mdm-siR277-1/2表达升高(J),而其靶基因MdRNL1/2/3/4/5的表达降低(K),叶片感病症状加重(L)、感病率升高(M)。
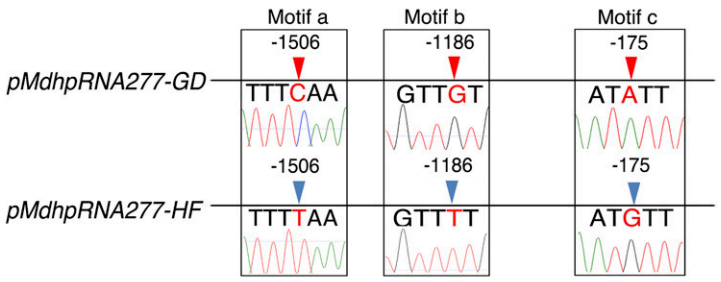
图14 GD和HF中MdhpRNA277启动子的三个SNP (Zhang et al., 2018)。作者克隆了MdhpRNA277的启动子序列(1711bp),并确定了GD和HF中 MdhpRNA277启动子的三个SNP(分别位于-1506、-1186和-175bp处)。

图15 pMdhpRNA277启动子区motif b对其活性至关重要 (Zhang et al., 2018)。作者为了确定哪个SNP对pMdhpRNA277的活性至关重要,将pMdhpRNA277-HF中的三个SNP(motif a、b和c)突变为GD序列,并将突变的启动子与GUS融合,发现当pMdhpRNA277-GD中的motif b突变为与pMdhpRNA277-HF中相同的序列(G至T)时,在接种ALT1后的GD和HF幼苗中均未检测到GUS染色(A、B)。
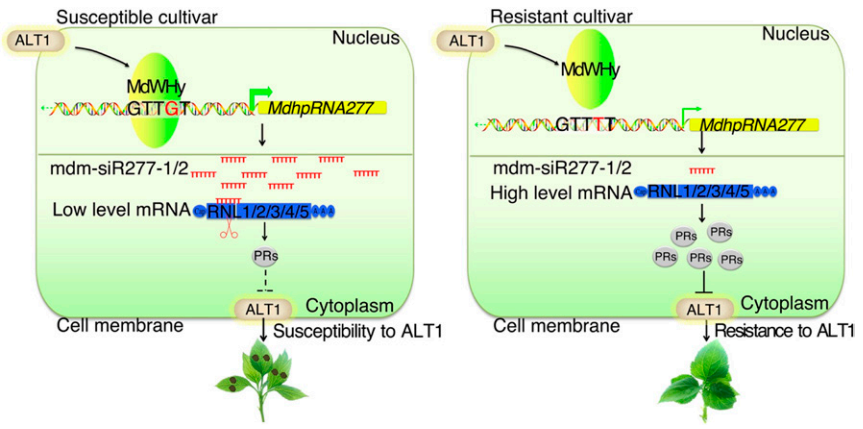
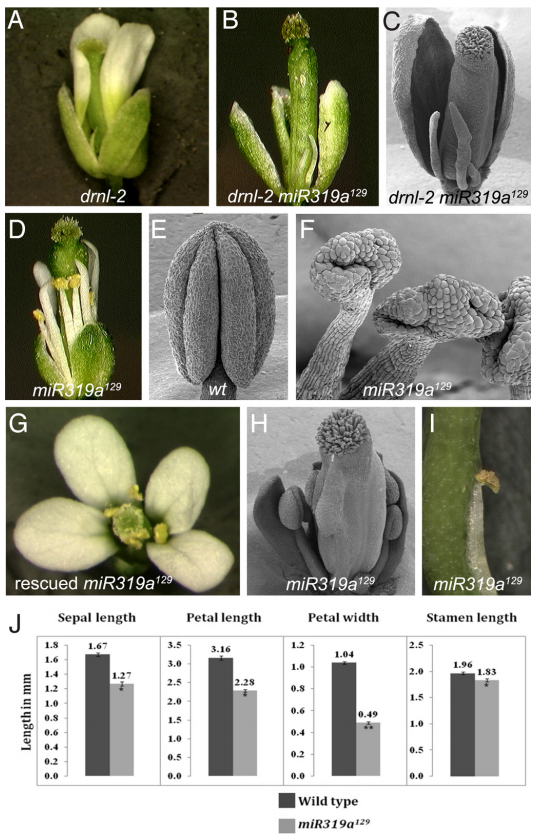
图17 miR319a129突变体在花发育中表现出明显的缺陷 (Nag et al., 2009)。drnl-2突变体花瓣长度不规则(A),drnl-2 miR319a129双突变体花瓣减少(B、C),miR319a129花药发育畸形(E、F),转化含有miR319a基因的3.7kb基因组DNA的T-DNA可恢复miR319a129突变体表型(G),与野生型相比miR319a129突变体雄蕊较短(D、H、I),与野生型相比miR319a129突变体花器官较小(J)。

图18 miR319a的单碱基突变会使其靶标基因TCP在花序中的表达增加 (Nag et al., 2009)。作者通过图位克隆的方法克隆了miR319a129基因,发现miR319a129突变体在成熟miR319a(At4g23713)的第12位含有G到A的单碱基突变(A),并且发现miR319a129功能缺失会使其靶标基因TCP在花序中的表达增加(B)。

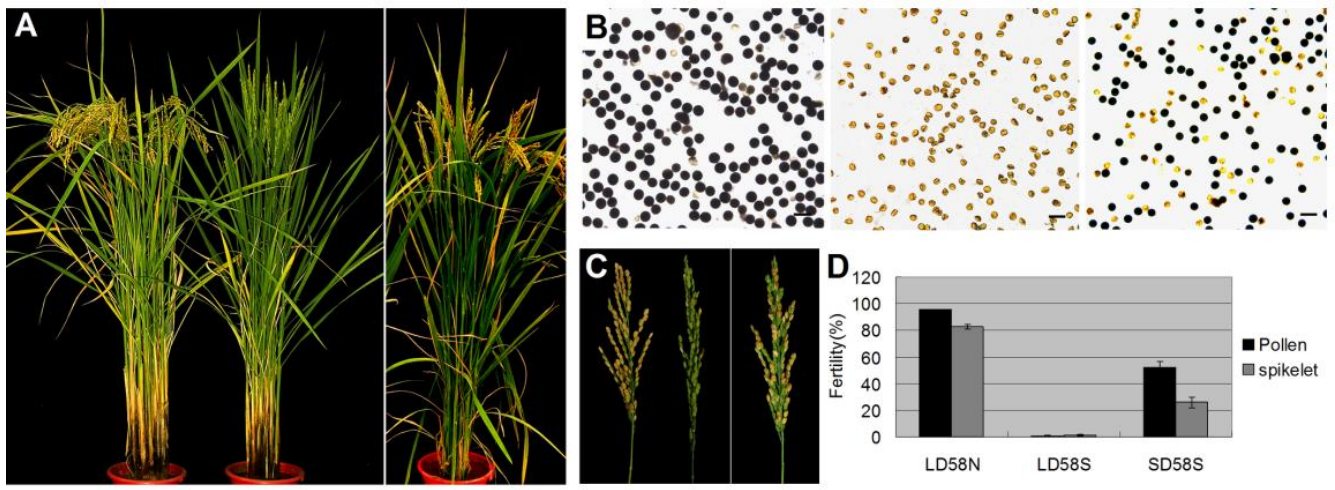
图20 58N和58S在自然长日照和短日照条件下的花粉育性 (Ding et al., 2012)。作者发现58N(左)在长日照条件下生长时花粉可育,58S(中)在长日照条件下生长时花粉完全不育,58S(右)在短日照条件下生长时花粉部分可育(A-D)。
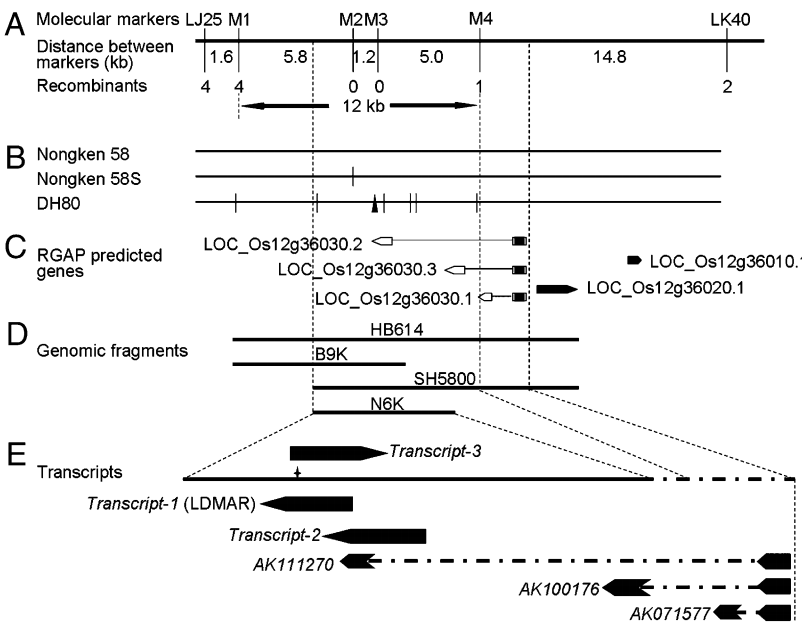
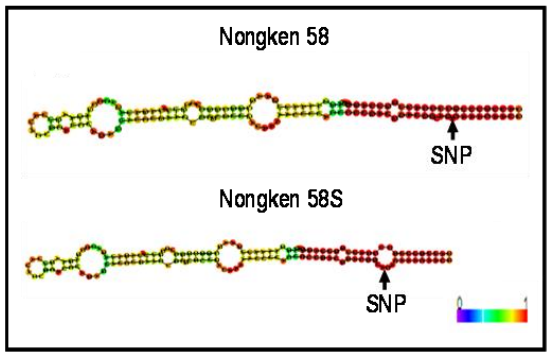
图21 pms3的图位克隆 (Ding et al., 2012)。已有文献表明pms3是将58N转变为58S的原始突变,作者通过比较58S、DH80和58N中28.4kb区域的测序发现一个由58N中的G替换58S中的C而产生的SNP(A-E)。
Chen Z, Ke W, He F, et al. A single nucleotide deletion in the third exon of FT-D1 increases the spikelet number and delays heading date in wheat (Triticum aestivum L.). Plant Biotechnol J. 2022;20(5):920-933. doi:10.1111/pbi.13773
Guo M, Yang F, Liu C, et al. A single-nucleotide polymorphism in WRKY33 promoter is associated with the cold sensitivity in cultivated tomato. New Phytol. 2022;236(3):989-1005. doi:10.1111/nph.18403
Fang J, Guo T, Xie Z, et al. The URL1-ROC5-TPL2 transcriptional repressor complex represses the ACL1 gene to modulate leaf rolling in rice. Plant Physiol. 2021;185(4):1722-1744. doi:10.1093/plphys/kiaa121.
Nag A, King S, Jack T. miR319a targeting of TCP4 is critical for petal growth and development in Arabidopsis. PNAS. 2009;106(52):22534-22539. doi:10.1073/pnas.0908718106.
Ding J, Lu Q, Ouyang Y, et al. A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice. PNAS. 2012;109(7):2654-2659. doi:10.1073/pnas.1121374109.
Huang Y, Chen X, Liu Y, et al. Mitochondrial AtPAM16 is required for plant survival and the negative regulation of plant immunity. Nat Commun. 2013;4:2558. doi:10.1038/ncomms3558
Zhang Q, Ma C, Zhang Y, et al. A Single-Nucleotide Polymorphism in the Promoter of a Hairpin RNA Contributes to Alternaria alternata Leaf Spot Resistance in Apple (Malus × domestica). Plant Cell. 2018;30(8):1924-1942. doi:10.1105/tpc.18.00042
