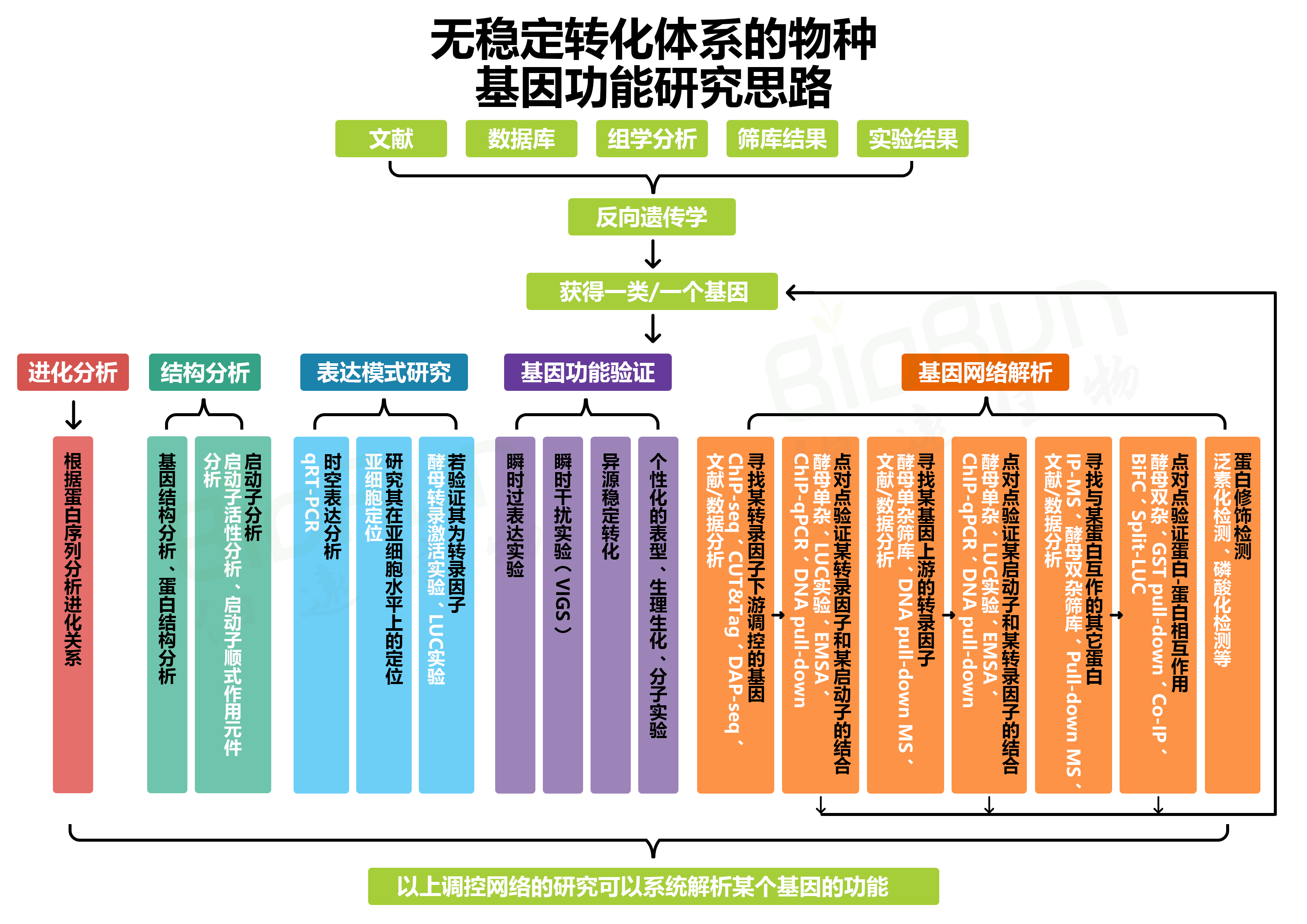
伯小远在“没有转化体系的物种,如何研究其基因功能(一)”中重点写了有稳定转化体系的物种的基因功能研究套路总结,在“(二)”中主要写了有哪些瞬时转化体系大家可以考虑使用,在“(三)”中主要写了无稳定转化体系的物种可以做的一些分子生物学实验、如何进行异源转化验证等。今天伯小远准备通过文献解读把前面写的知识点给串成一个相对完整的套路总结,我们来一起看看吧。本次推文通过文献最终总结出来的研究思路如下:
作者通过前期的全基因组分析及相关实验得到了一个与干旱有关的基因RcNAC091,先对其进行系统发育分析(图1)。
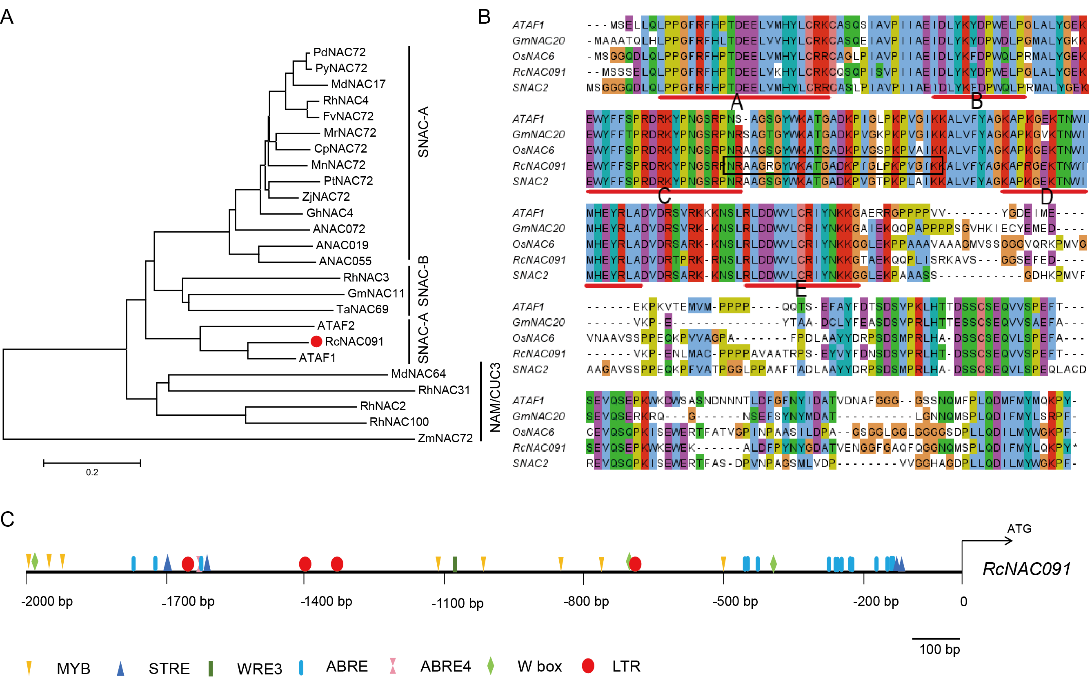
图1 RcNAC091进化关系分析 (Geng et al., 2023)。
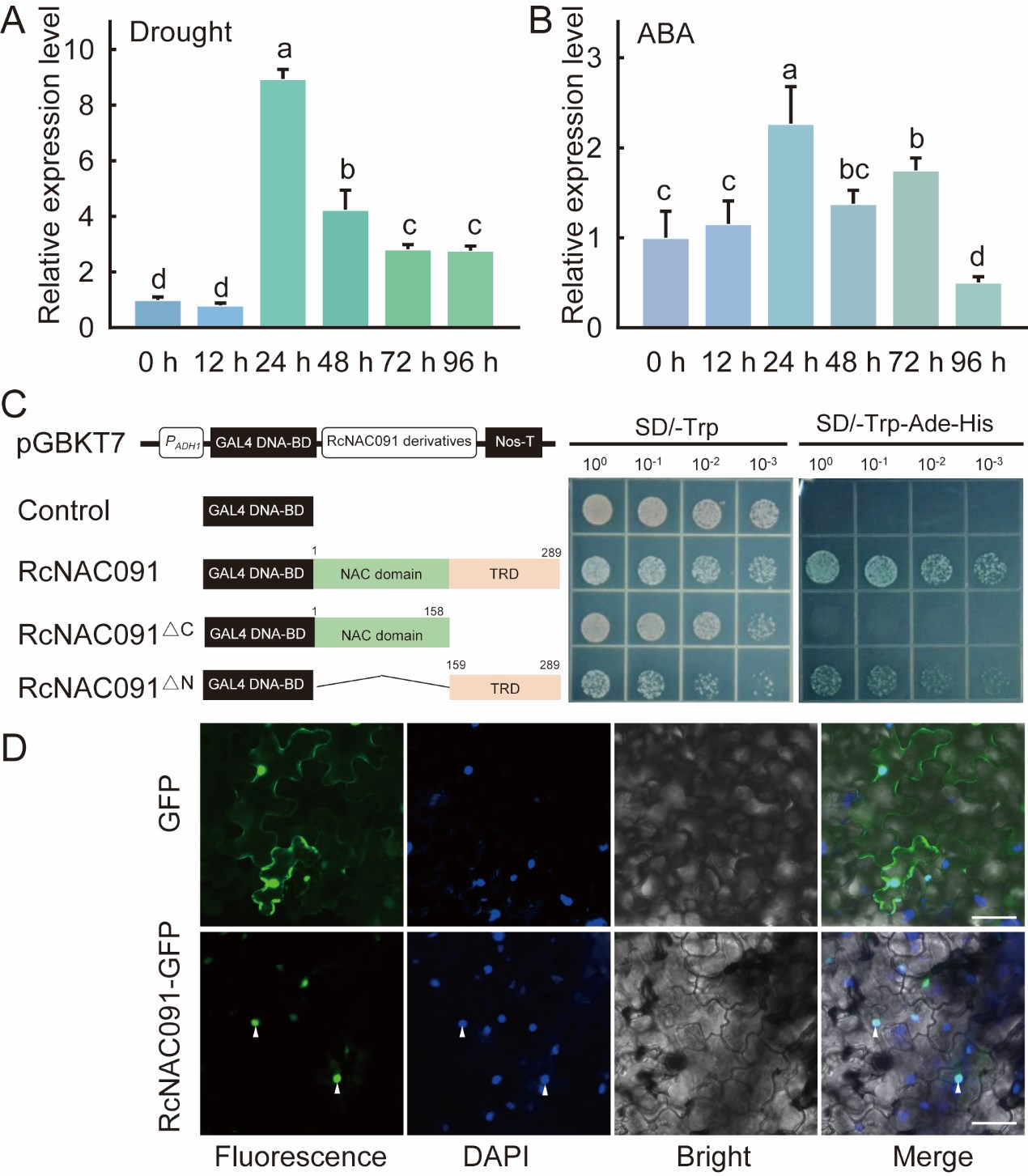
备注:关于图2C、图2D实验的具体内容,可参考文章“最强大脑炼成记——如何验证一个基因是转录因子”。
对RcNAC091基因的功能研究:在月季叶片中进行瞬时过表达实验及相关检测(图3A-E)、在月季植株上进行病毒诱导的基因沉默实验(VIGS)及相关检测(图3F-K)。

图3 (A)脱水处理0、4h、8h后再复水3h,观察对照叶片(VC)和过表达RcNAC091基因的叶片(瞬转入质粒pSuper:RcNAC091)的表型,结果显示,相比于对照组,过表达RcNAC091基因的叶片卷曲程度更低;(B)定量PCR检测(A)中两种材料中RcNAC091基因的相对表达量;(C-E)对(A)中的两种材料进行鲜重、叶绿素含量、电解质泄露率的测定。结果显示,过表达RcNAC091基因的叶片鲜重更高、叶绿素含量更高、电解质泄露率更低;(F)用20% PEG 6000处理0、2d、4d后再恢复浇水3d,观察对照植株(TRV)和VIGS植株(TRV-RcNAC091)的表型;(G)定量PCR检测(F)中两种材料中RcNAC091基因的相对表达量;(H-I)对(F)中的两种材料进行叶绿素含量测定、电解质泄露率测定,结果显示,相比于对照组,TRV-RcNAC091植株叶绿素含量更低、电解质泄露率更高;(J-K)20% PEG 6000处理两天后,对TRV和TRV-RcNAC091叶片进行台盼蓝染色、NBT染色、DAB染色、O2含量测定,结果显示,TRV-RcNAC091植株细胞死亡变多、ROS含量变高、O2含量更高。以上结果表明过表达RcNAC091会提高月季的耐旱性,沉默RcNAC091会降低月季的耐旱性 (Geng et al., 2023)。
探索ABA、RcNAC091、干旱之间的联系(图4)。
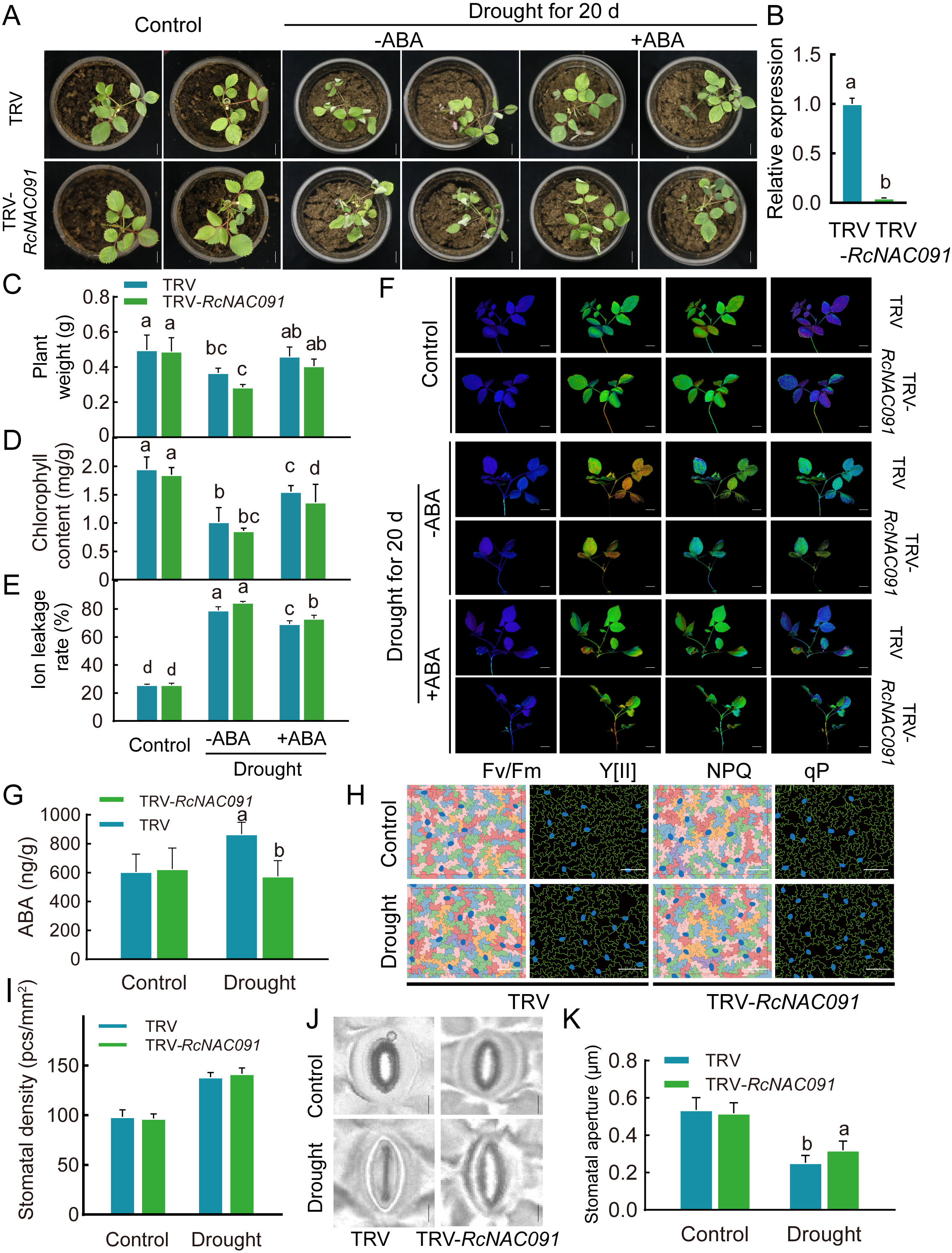
图4 ABA介导了由RcNAC091调控的耐旱性。方法:干旱处理20d且使用ABA处理或不处理TRV-RcNAC091植株。可以观察到经干旱处理后,与对照TRV相比,TRV-RcNAC091植株更小、更枯萎、叶子偏黄(A),其植株重量更低(C)、叶绿素含量更低(D)、电解质泄露率更高(E),但经ABA处理后,发现ABA可以增加植株重量(C)、增加叶绿素含量(D)、降低电解质泄露率(E)。同时,叶绿素荧光参数的测定结果也表明ABA能提高植株的耐旱性(F)。经干旱处理后,与对照TRV相比,TRV-RcNAC091植株中的ABA含量更低(G)、气孔孔径更大(J、K)。以上结果表明,ABA能增强植株耐旱性,RcNAC091以ABA介导的方式提高了植株耐旱性 (Geng et al., 2023)。
通过转录组寻找RcNAC091调控的下游基因(图5)。
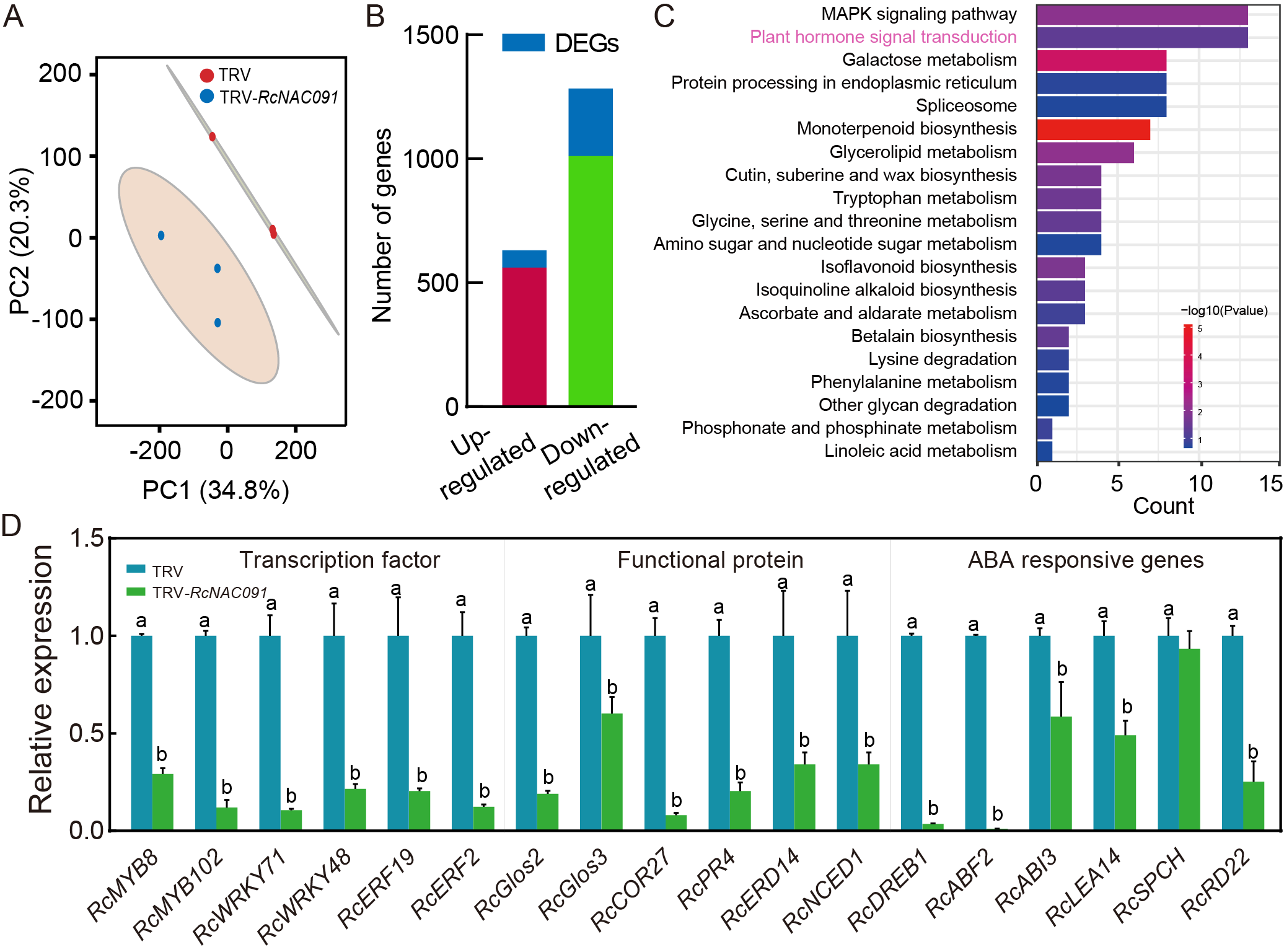
图5 通过鉴定TRV和TRV-RcNAC091的差异表达基因,最终查找了已知的参与干旱响应的18个基因(D),并将其中的RcWRKY71和RcERF2两个基因作为RcNAC091的候选下游基因 (Geng et al., 2023)。
检测RcNAC091是否结合RcWRKY71的启动子(图6)。
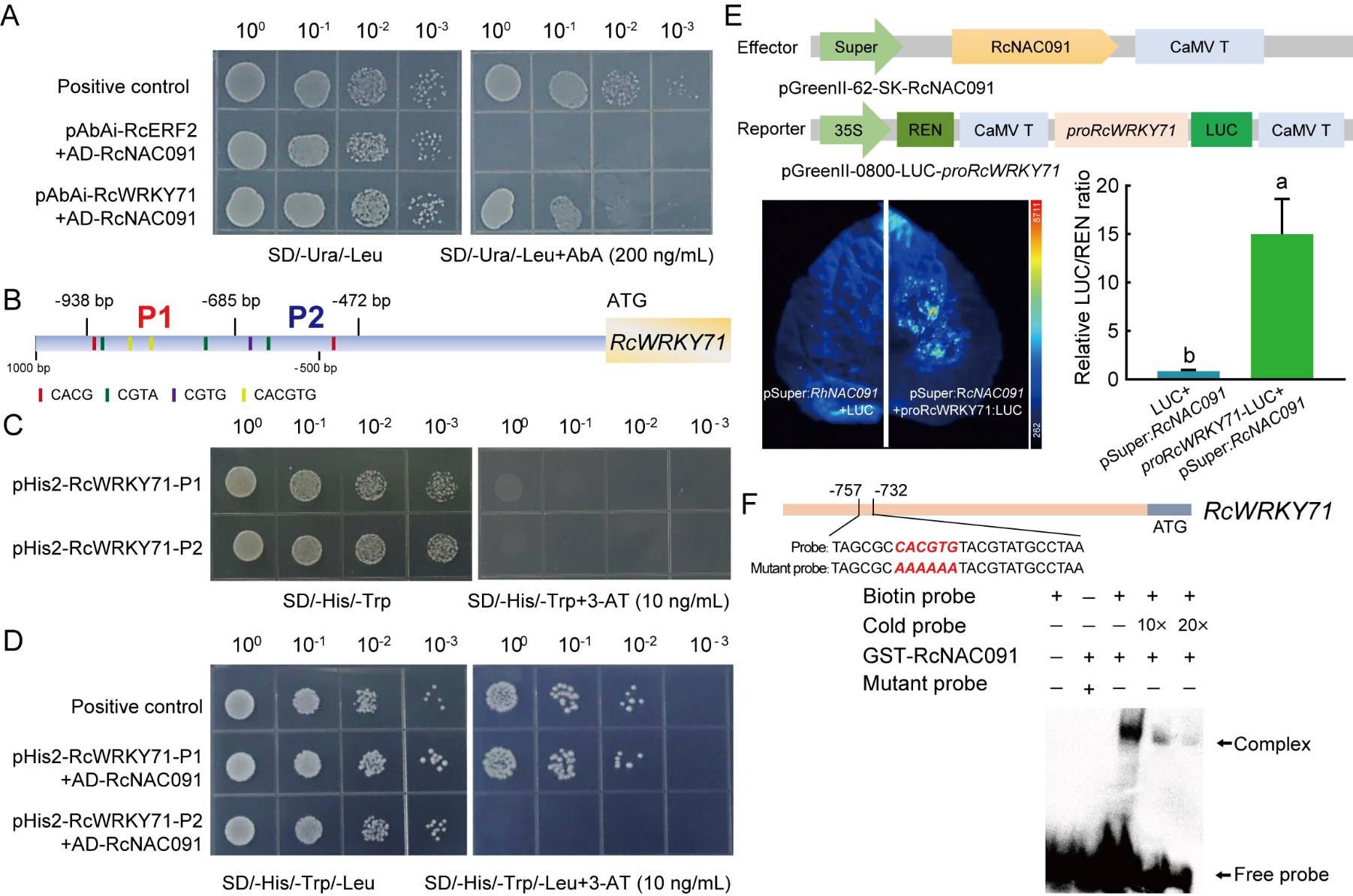
图6 通过酵母单杂交实验(A-D)、双荧光素酶报告基因实验(E)、EMSA实验(F)证明RcNAC091可结合RcWRKY71的启动子。具体来说,是结合RcWRKY71启动子P1区内的CACGTG元件 (Geng et al., 2023)。
备注:关于酵母单杂交实验,可参考文章“酵母杂交那些事儿(一)”。关于双荧光素酶报告基因实验,可参考文章“天呐!这个实验系统也太强大了吧”。关于EMSA实验,可参考文章“蛋白-核酸相互作用之EMSA实验正式上线啦”。
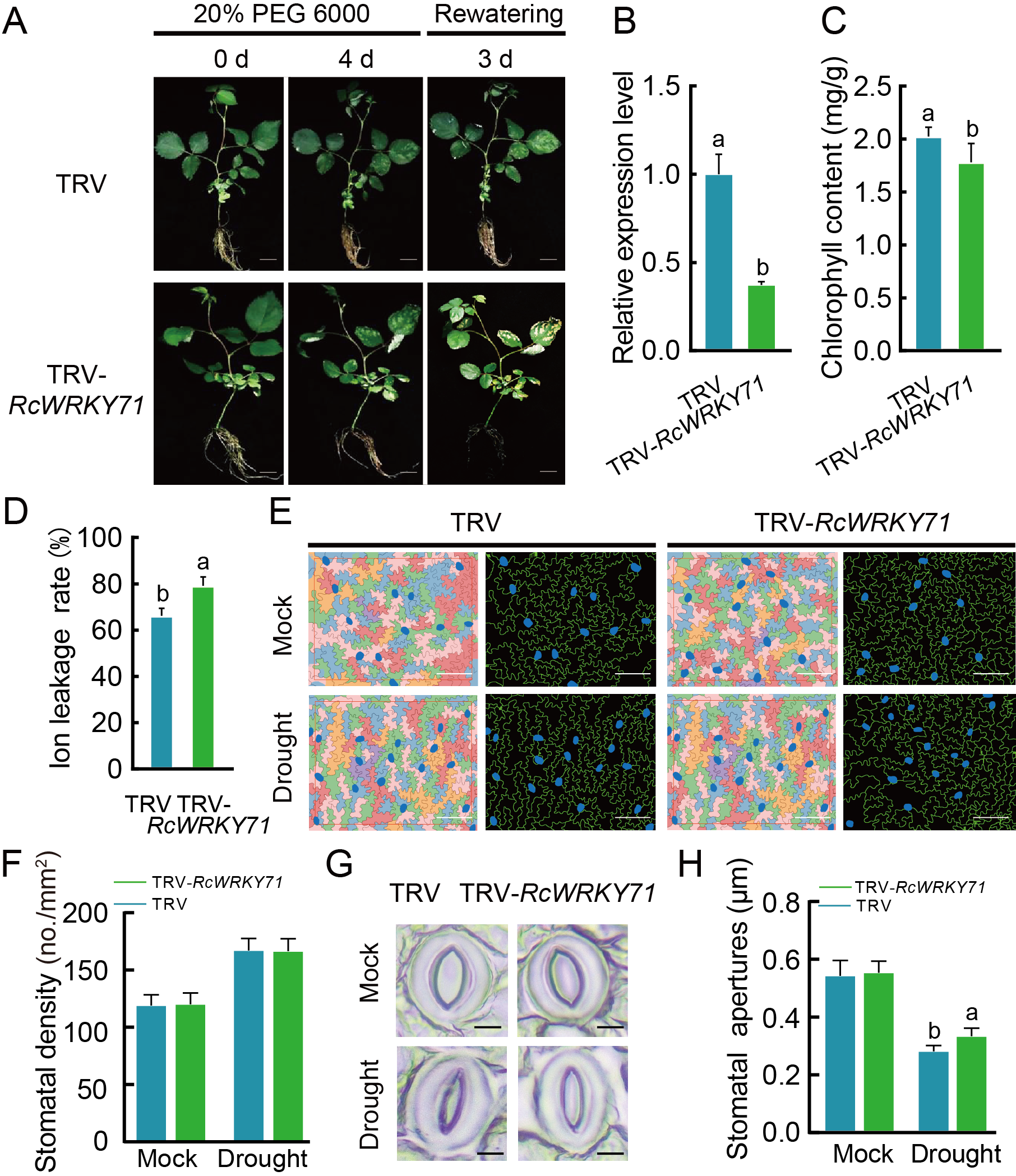
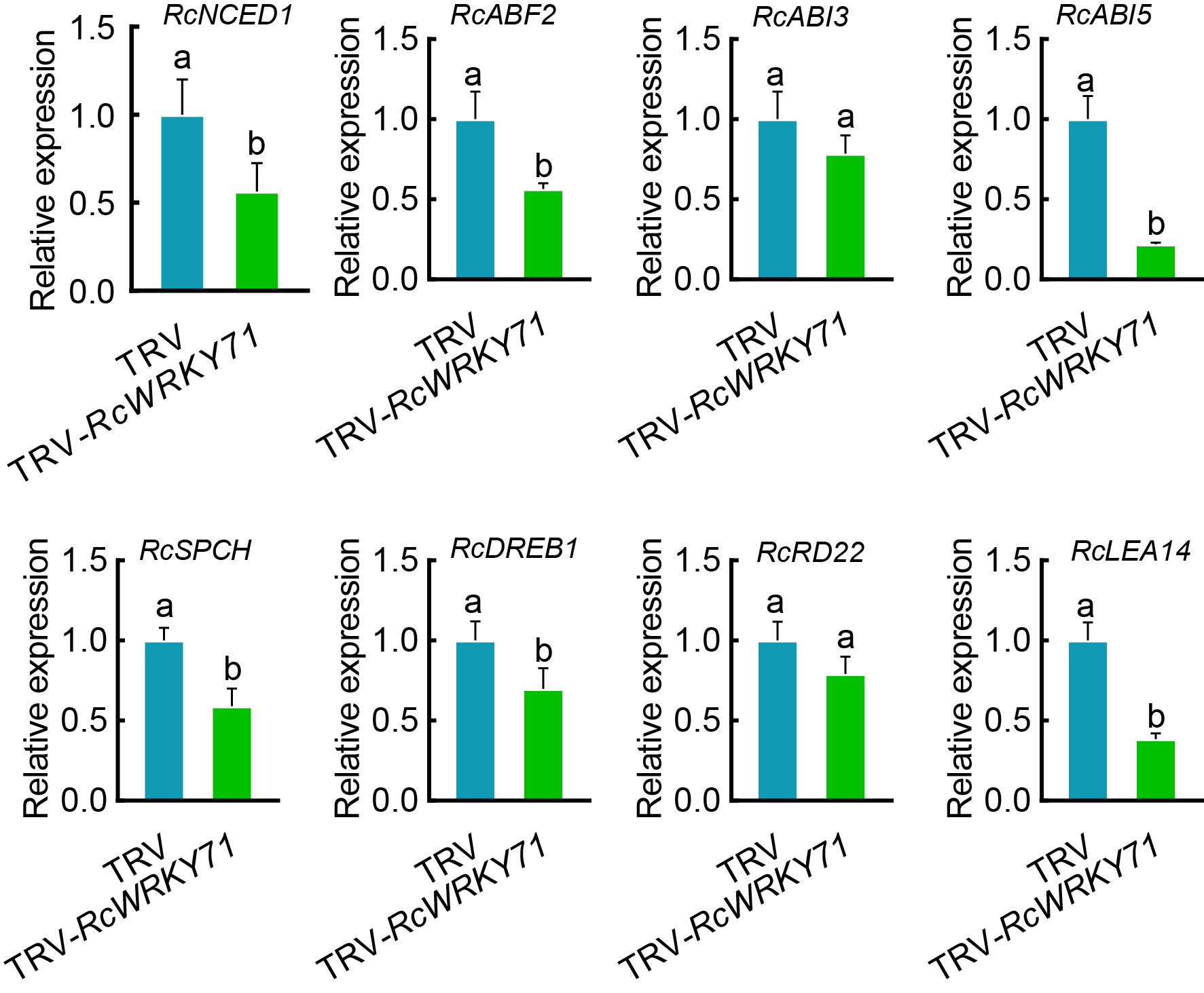
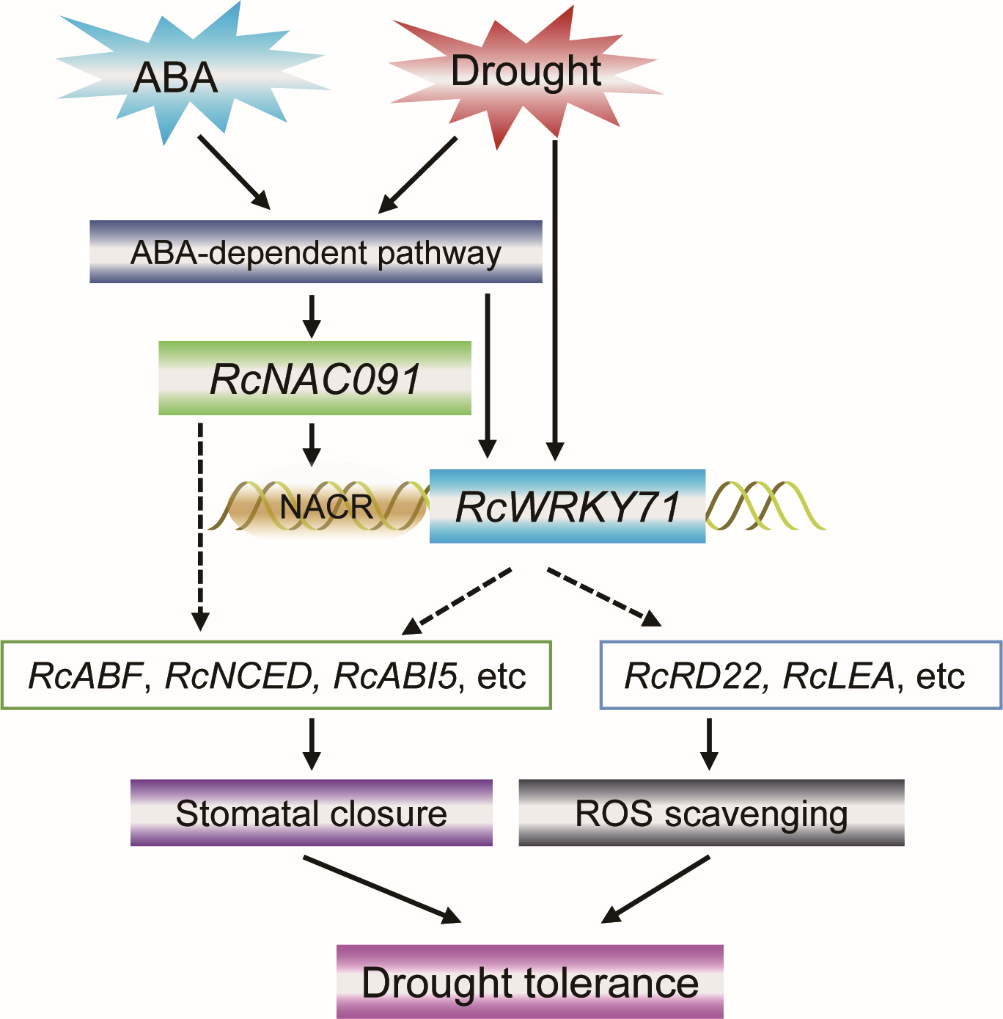
探索ABA、RcWRKY71、干旱之间的联系(图8)。
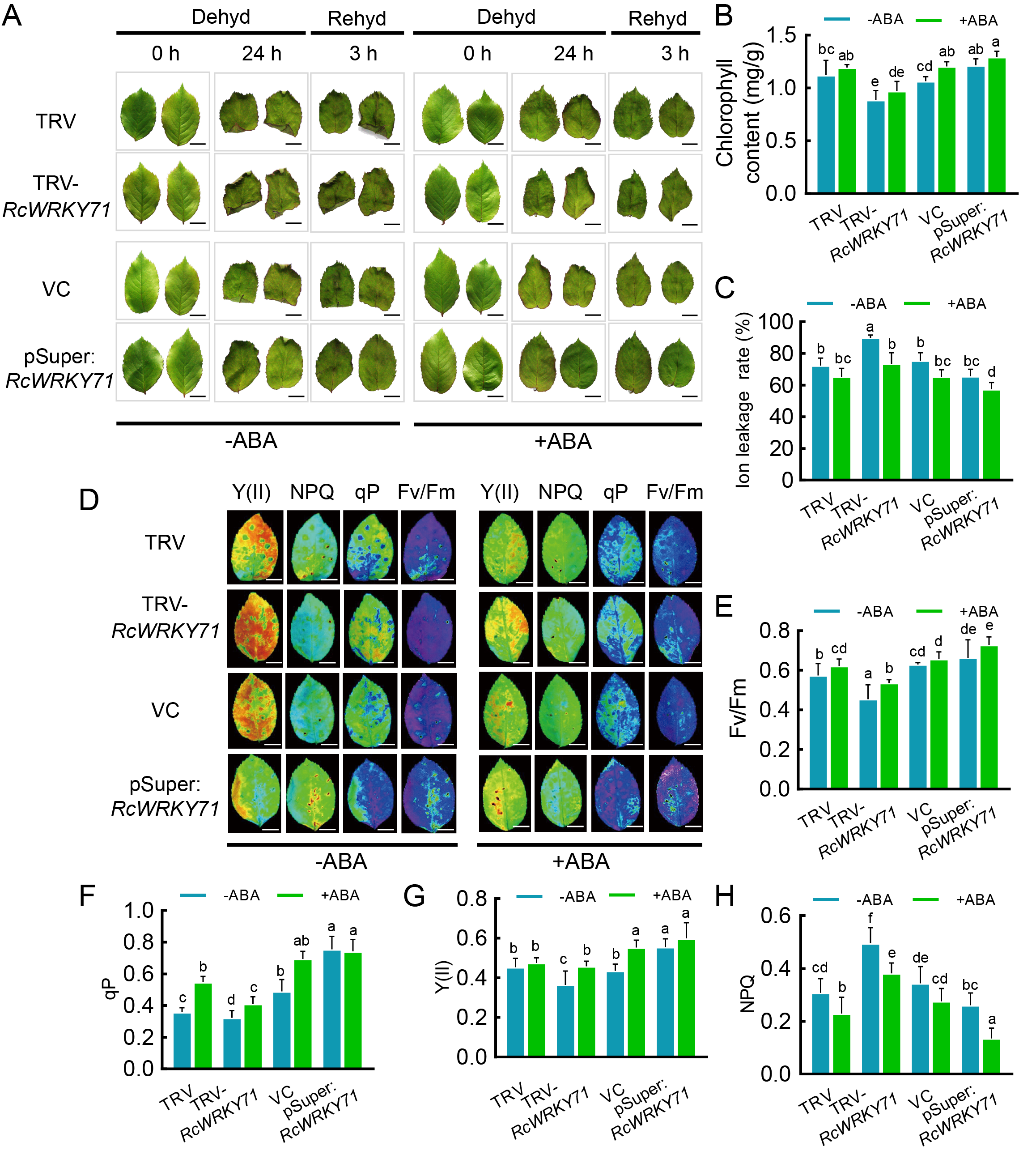
图8 ABA介导了由RcWRKY71调控的耐旱性。在月季叶片中进行VIGS和过表达实验,干旱条件下,经ABA处理或不处理,再进行相关检测,以上实验证明RcWRKY71正向调控由ABA介导的耐旱性 (Geng et al., 2023)。
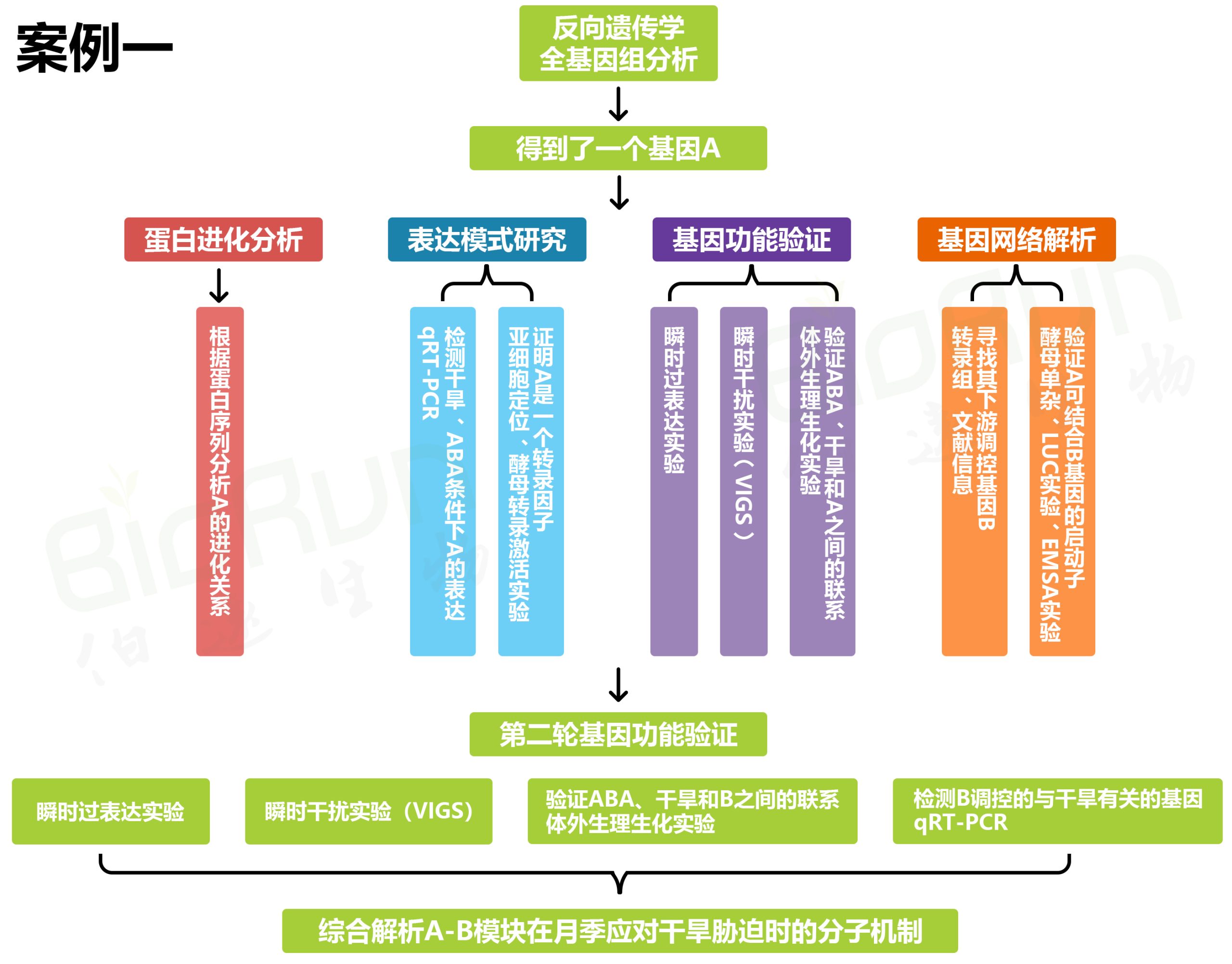
伯小远将以上研究思路整理如下:
首先,通过反向遗传学鉴定出候选lncRNA109897及其可能的靶基因JrCCR4(图11)。
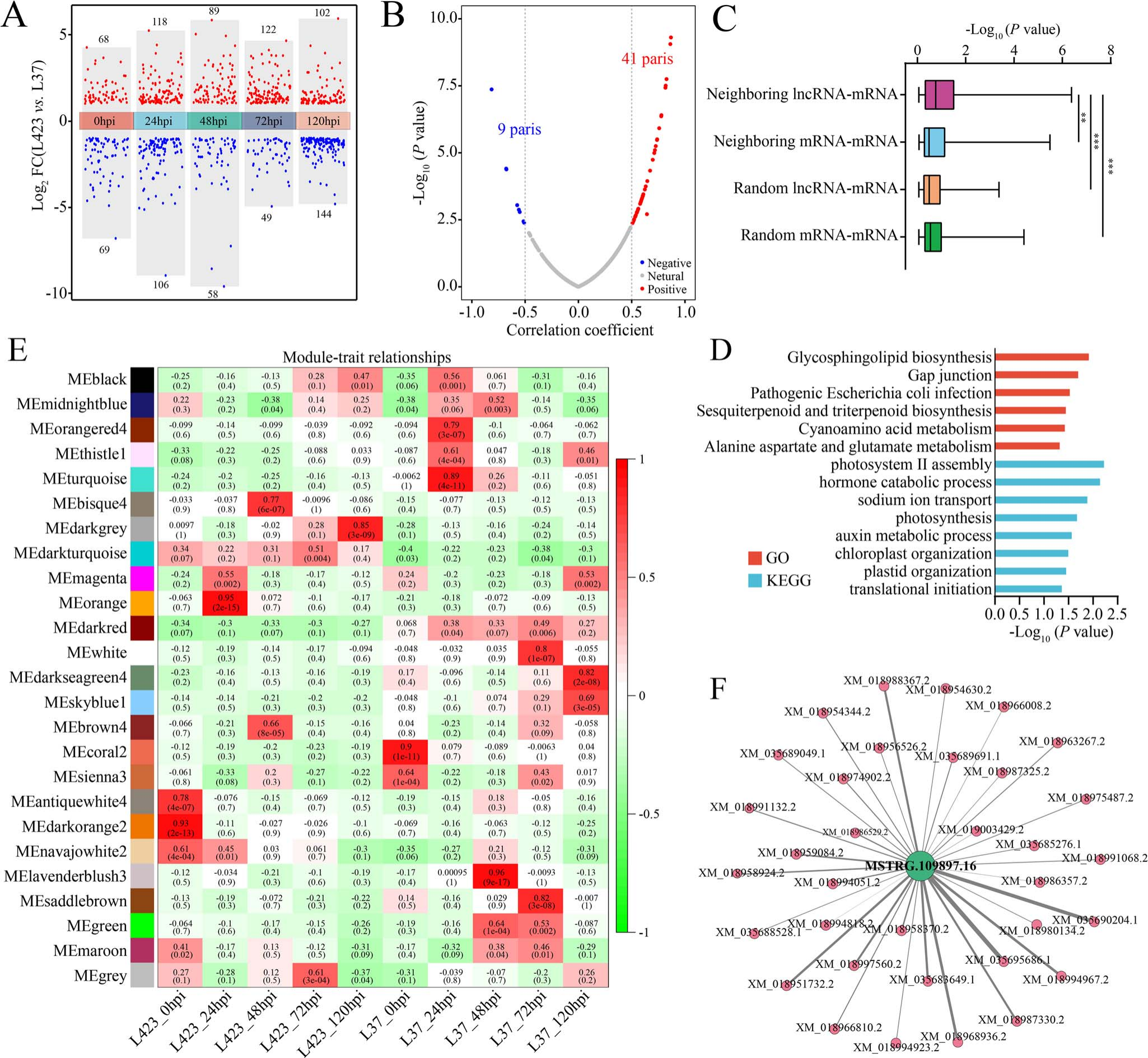
图11 通过全基因组鉴定核桃中对炭疽病作出响应的lncRNAs,对差异表达的lncRNAs进行分析,再通过加权基因共表达网络分析(WGCNA)得出了与核桃抗炭疽病显著相关的中心lncRNA即MSTRG.109897.16(lncRNA109897)(F),在与其连接的基因中,XM_035695686.1(JrCCR4)和lncRNA109897之间可能存在正向的调控关系,同时,JrCCR4也是lncRNA109897的顺式靶标(B) (Zhou et al., 2023)。
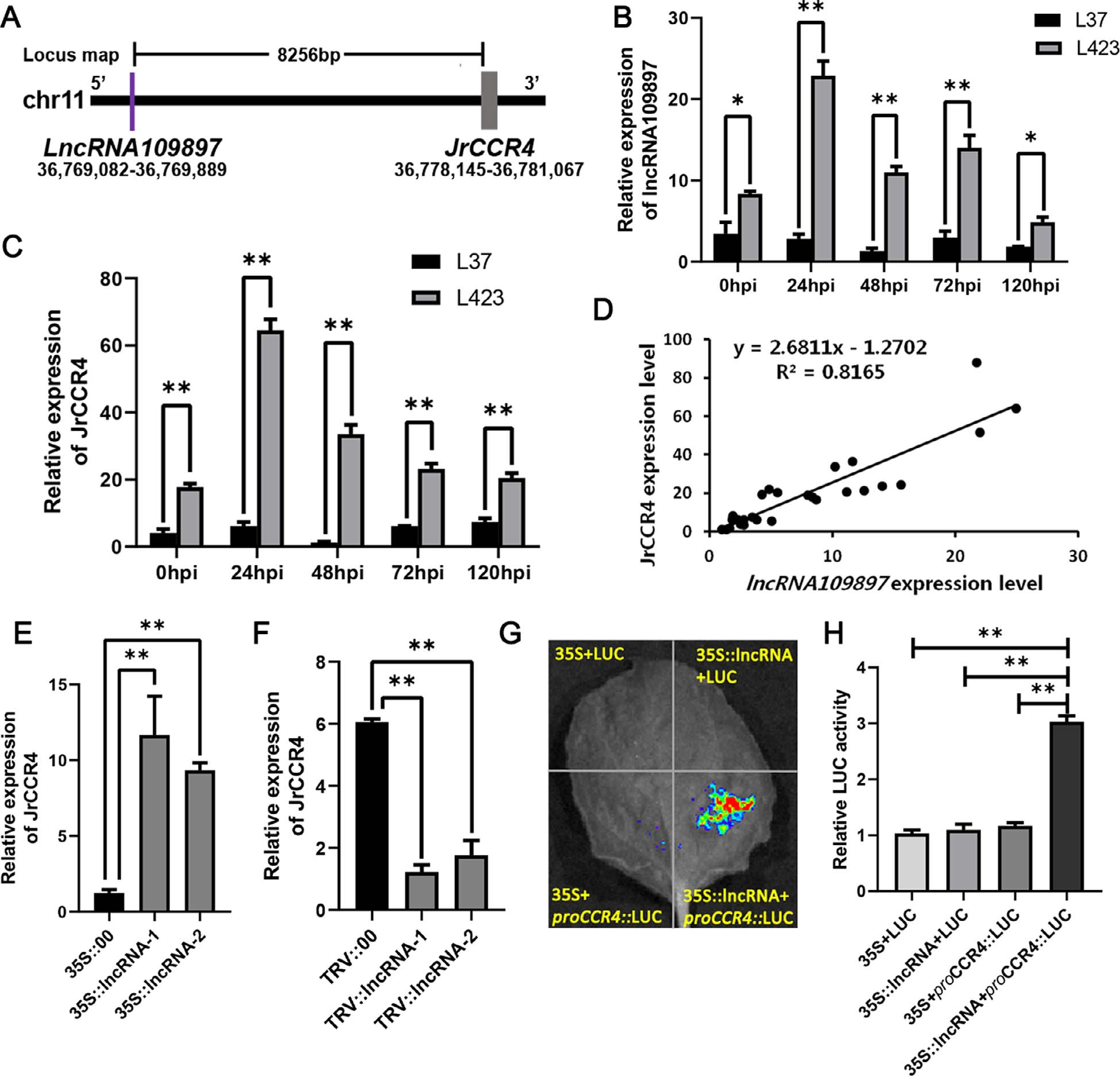
对lncRNA109897和JrCCR4基因进行功能研究:在核桃叶片中进行瞬时过表达实验、VIGS实验及相关检测(图13)。
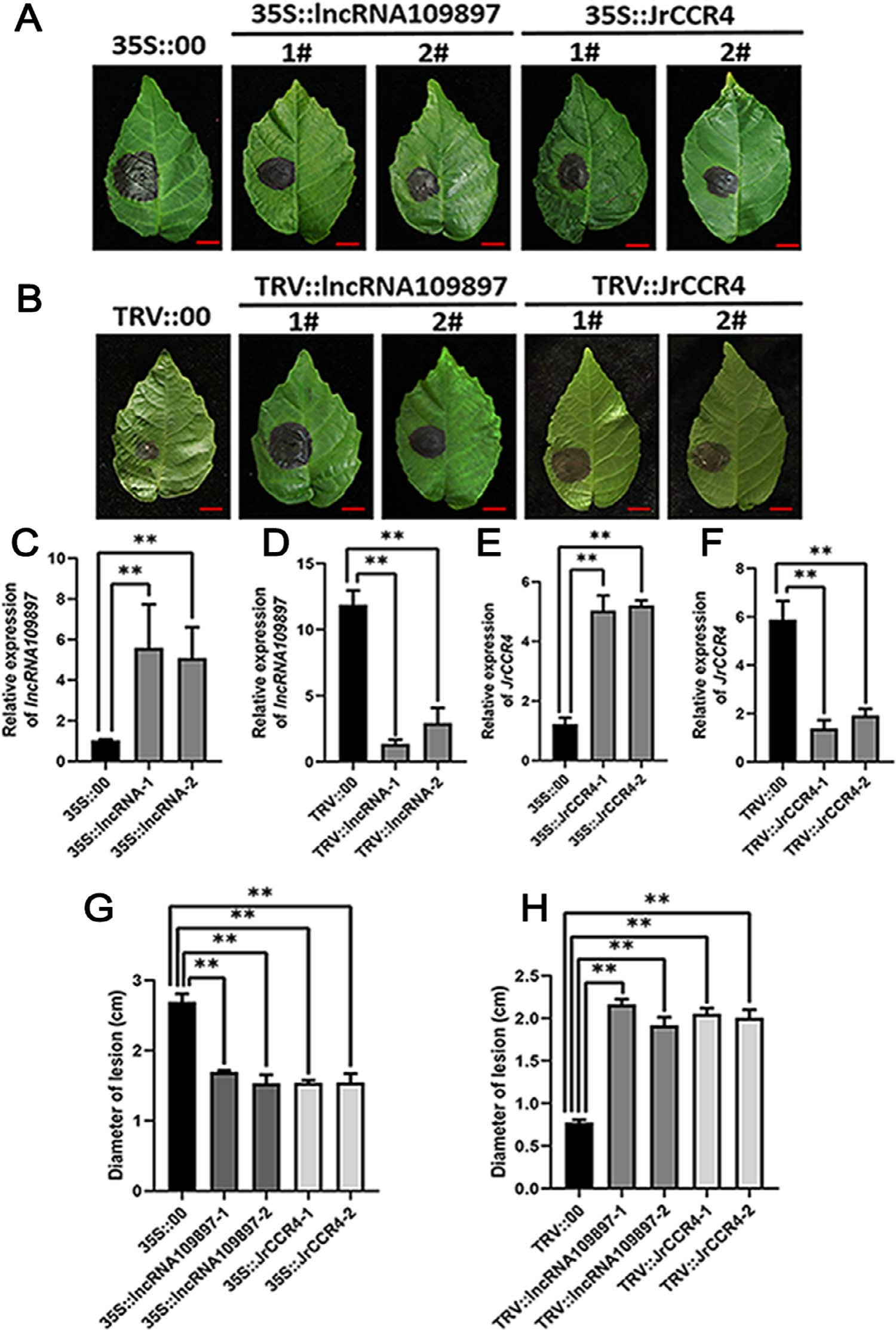
图13 结果显示,接种了炭疽病菌的35S::lncRNA109897、35S::JrCCR4的核桃叶片比对照的病斑更小,而相反,接种了炭疽病菌的TRV::lncRNA109897、TRV::JrCCR4的核桃叶片比对照的病斑更大(A、B)。另外,作者还在拟南芥中进行了稳定的过表达JrCCR4基因的实验,发现其对炭疽病的抗性高于野生型(在原文的补充材料中) (Zhou et al., 2023)。
JrCCR4的亚细胞定位实验、寻找与JrCCR4相互作用的蛋白并进行验证(图14)。

图14 结果显示,JrCCR4定位于细胞核和细胞膜(A),以JrCCR4为诱饵进行酵母双杂筛库,得到了其潜在的相互作用蛋白JrTLP1b,再用酵母双杂交实验、GST pull-down、BiFC实验证明JrCCR4和JrTLP1b确实存在相互作用 (Zhou et al., 2023)。
备注:关于酵母双杂实验,可参考文章“酵母杂交那些事儿(三)”,关于GST pull-down实验,可参考文章“蛋白互作研究方法之GST pull-down/GST pull-down MS技术”,关于BiFC实验,可参考文章“BiFC实验protocol”。
检测JrCCR4和JrTLP1b启动子的关系(图15)。
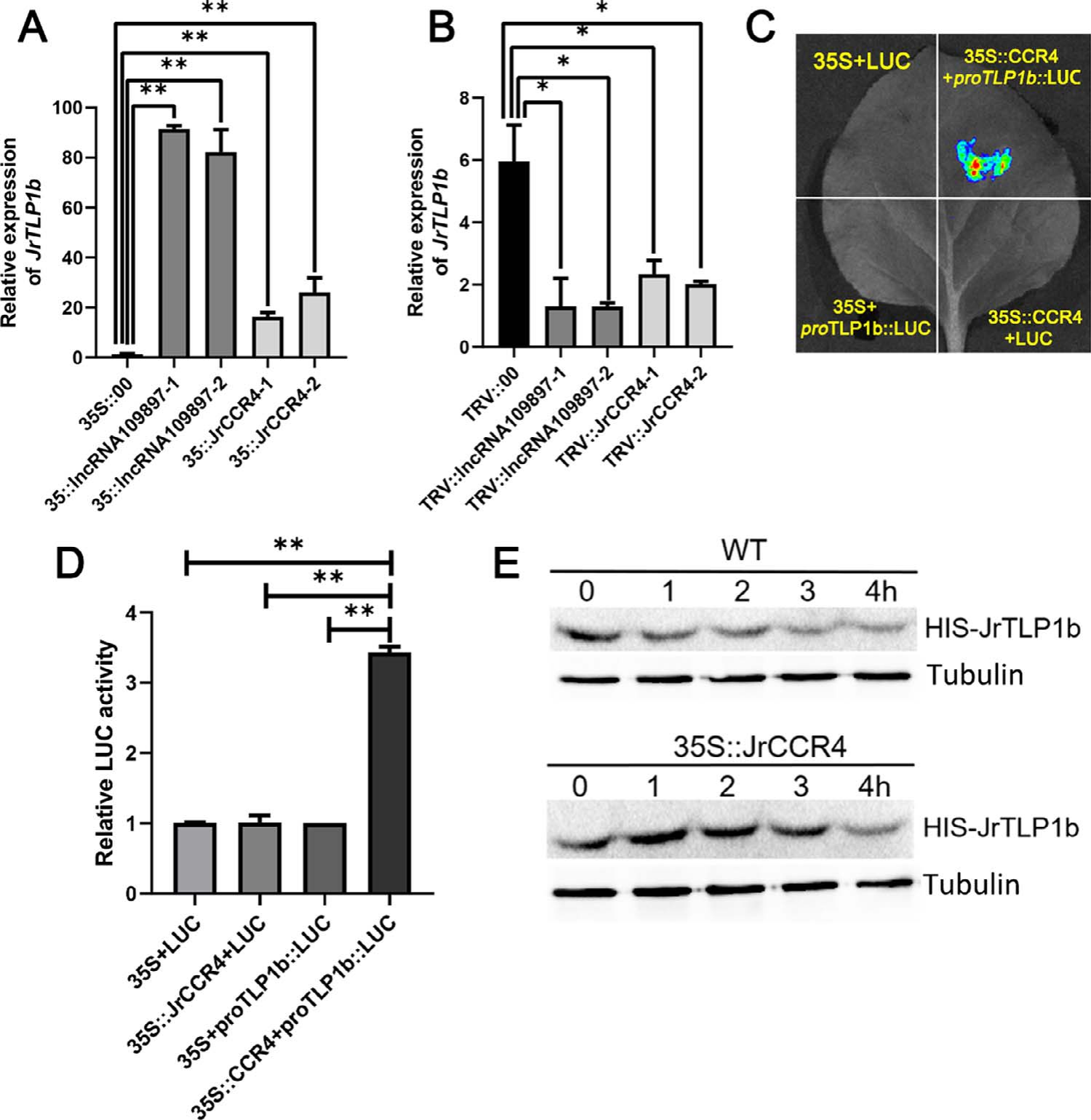
图15 结果显示,双荧光素酶报告基因实验(C、D)发现JrCCR4可结合JrTLP1b的启动子,体外实验表明JrCCR4并不会影响JrTLP1b的蛋白稳定性甚至增强了其稳定性(E) (Zhou et al., 2023)。
对JrTLP1b基因的功能研究:在核桃叶片中进行瞬时过表达实验、VIGS实验及相关检测;检测JrTLP1b和lncRNA109897启动子的关系(图16)。
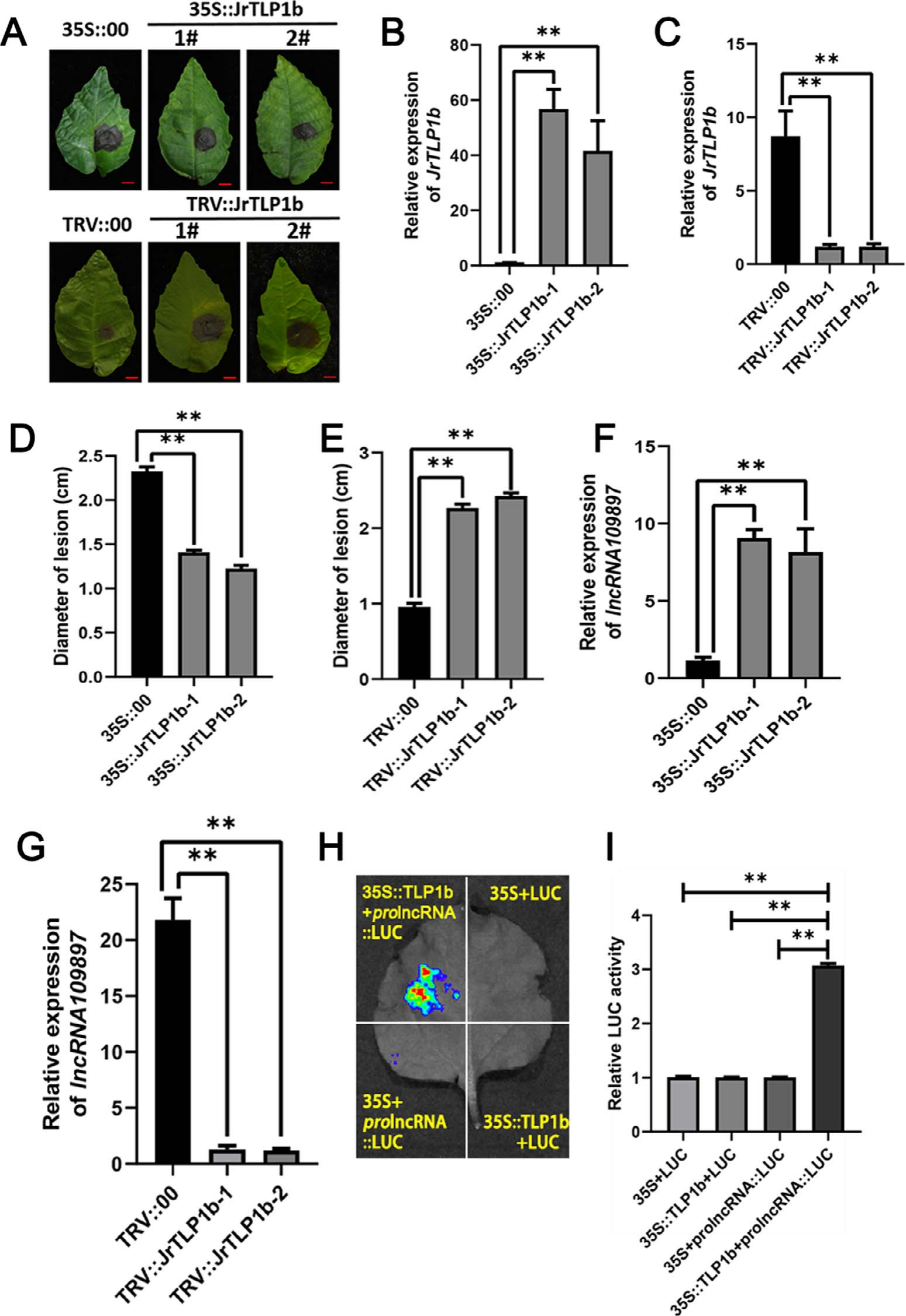
图16 结果显示,接种了炭疽病菌的35S::JrTLP1b的核桃叶片病斑比对照的病斑更小(A、D)、接种了炭疽病菌的TRV::JrTLP1b的核桃叶片病斑比对照的病斑更大(A、E)。双荧光素酶报告基因实验(H、I)发现JrTLP1b可结合lncRNA109897的启动子 (Zhou et al., 2023)。
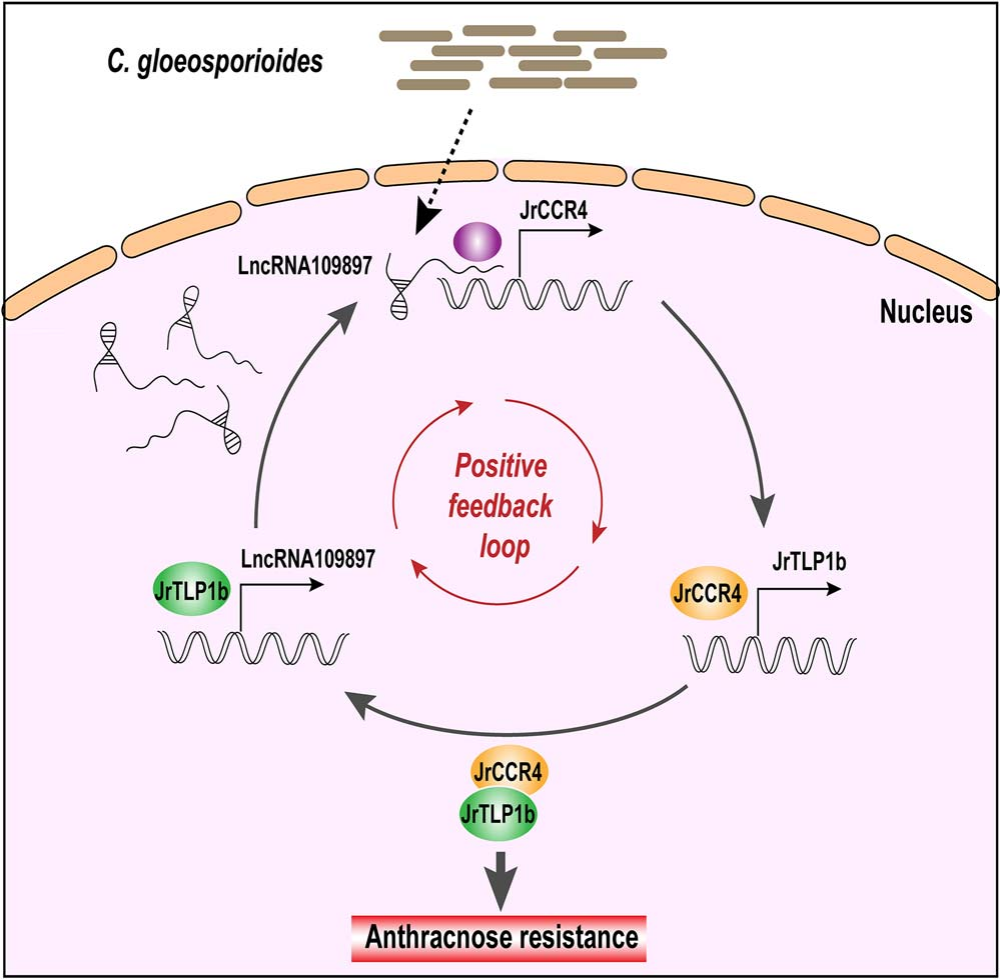
图17 lncRNA109897-JrCCR4-JrTLP1b在核桃抗炭疽病菌时的工作模型 (Zhou et al., 2023)。作者提供了lncRNA109897-JrCCR4-JrTLP1b正反馈调节核桃炭疽病菌抗性的证据,lncRNA109897正向影响其靶标JrCCR4,通过相互作用增加JrTLP1b的表达,而JrTLP1b又导致lncRNA109897表达增加,该调控通路的鉴定有助于进一步了解核桃的抗病机制。
伯小远将以上研究思路整理如下:
伯小远根据以上这些文献,再结合之前的经验整理了一个无稳定转化体系的物种基因功能研究思路:
2023年,西北农林科技大学张新梅、康振生课题组在Horticulture Research杂志上发表了题为“CaREM1.4 interacts with CaRIN4 to regulate Ralstonia solanacearum tolerance by triggering cell death in pepper”的文章,作者主要研究了辣椒中CaREM1.4在抗青枯病菌中的正向调控作用,后又通过实验证明了CaREM1.4与CaRIN4-12相互作用,对CaRIN4-12的基因功能研究证明了其在辣椒抗青枯菌过程中的负调控作用。
作者使用REM C端的保守序列搜索了辣椒的基因组序列,鉴定出了18个CaREM基因,并与拟南芥、番茄和水稻中的REM成员构建了发育进化树(图18)。
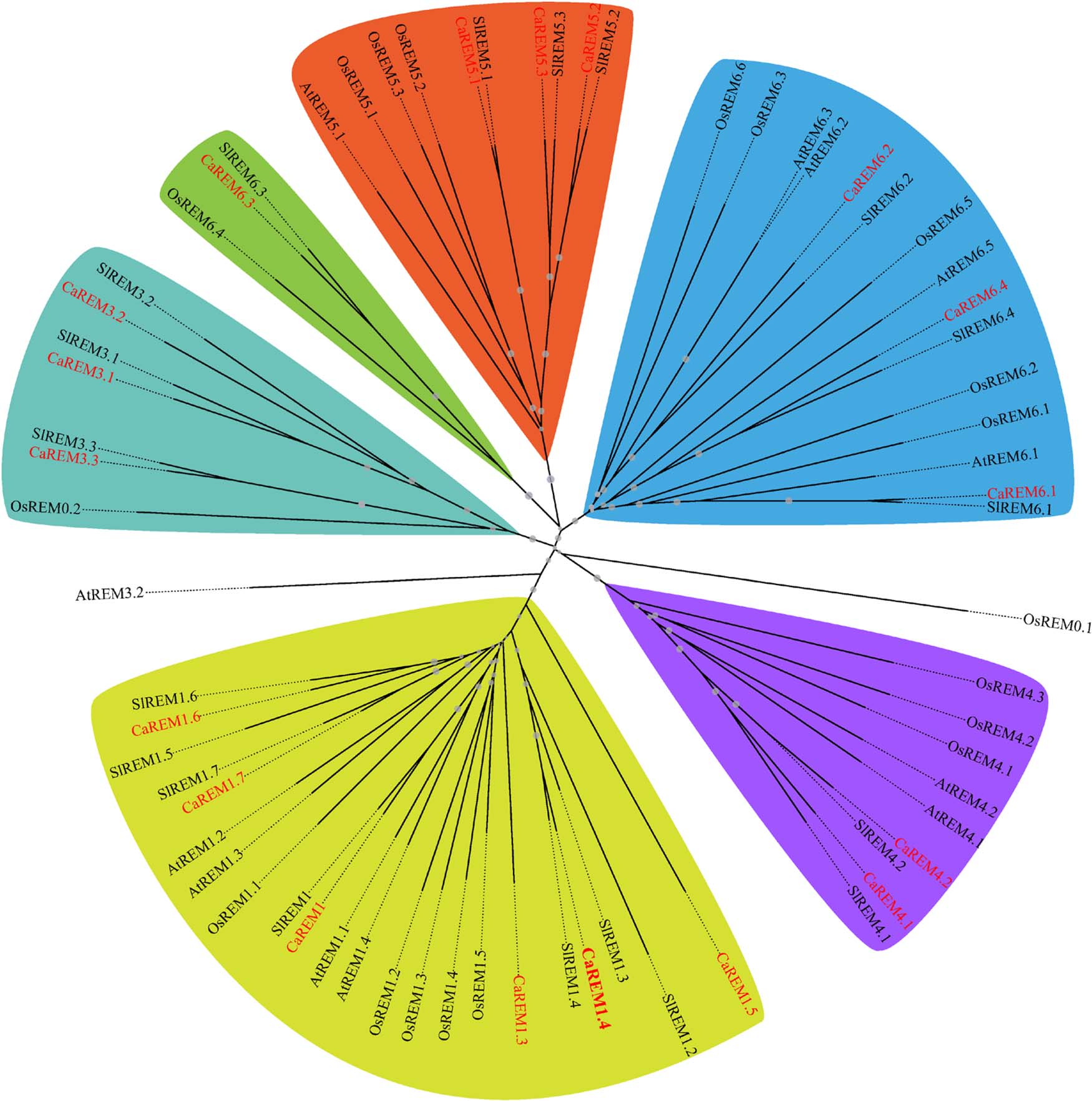
图18 进化分析 (Zhang et al., 2023)。
对18个CaREM进行基因结构分析(见原文表格中)、蛋白保守结构域分析(图19)以及启动子顺式作用元件分析(见原文补充数据中)。
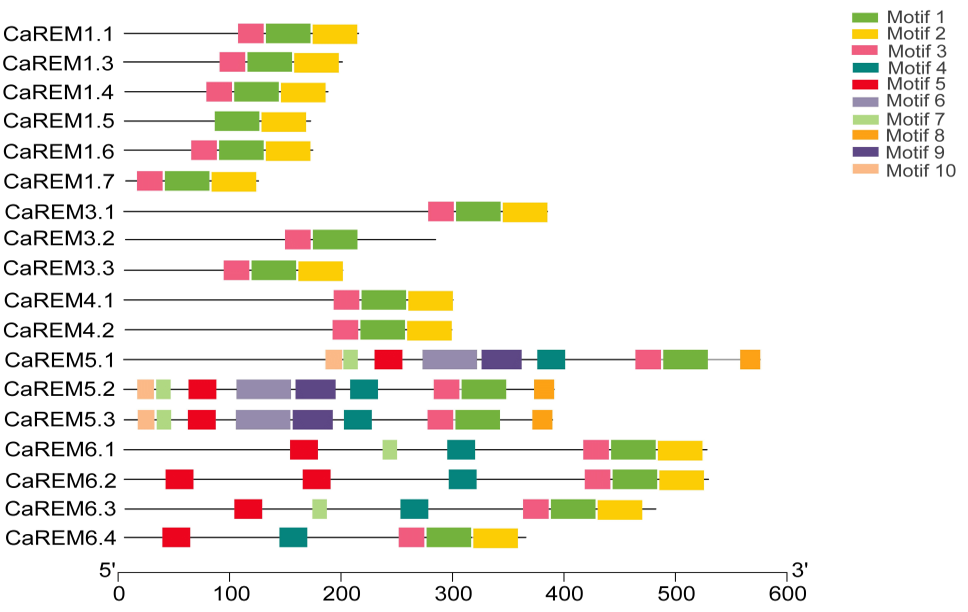
图19 辣椒中18个REM的蛋白保守基序示意图 (Zhang et al., 2023)。
对接种了青枯菌的辣椒进行CaREM基因的表达模式分析,发现CaREM1.4基因对青枯菌的反应最为显著(图20)。因此作者在后面的文章中重点研究了CaREM1.4基因在辣椒受青枯菌胁迫下所发挥的作用。
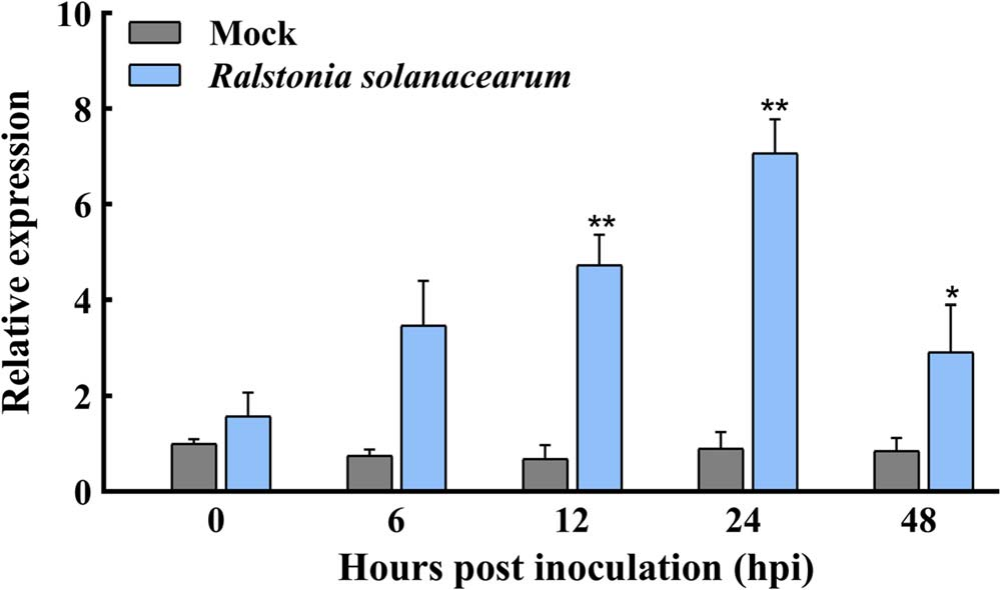
图20 CaREM1.4基因在受到青枯菌胁迫下的诱导表达 (Zhang et al., 2023)。
接着,作者对CaREM1.4进行基因功能研究,包括VIGS以及表型、疾病指数、青枯菌生长情况、各CaREM基因的转录水平检测、各免疫相关基因的转录水平检测(图21),瞬时过表达以及表型、相关生理生化实验、各免疫相关基因的转录水平检测(图22)。作者也检测了CaREM1.4的C端、N端及完整蛋白的亚细胞定位情况,以及其C端、N端在抗病中的作用(见原文补充数据中)。
以上实验表明,CaREM1.4为辣椒与青枯菌互作的正调控因子,并且其C端是引起细胞死亡的主要功能结构域。
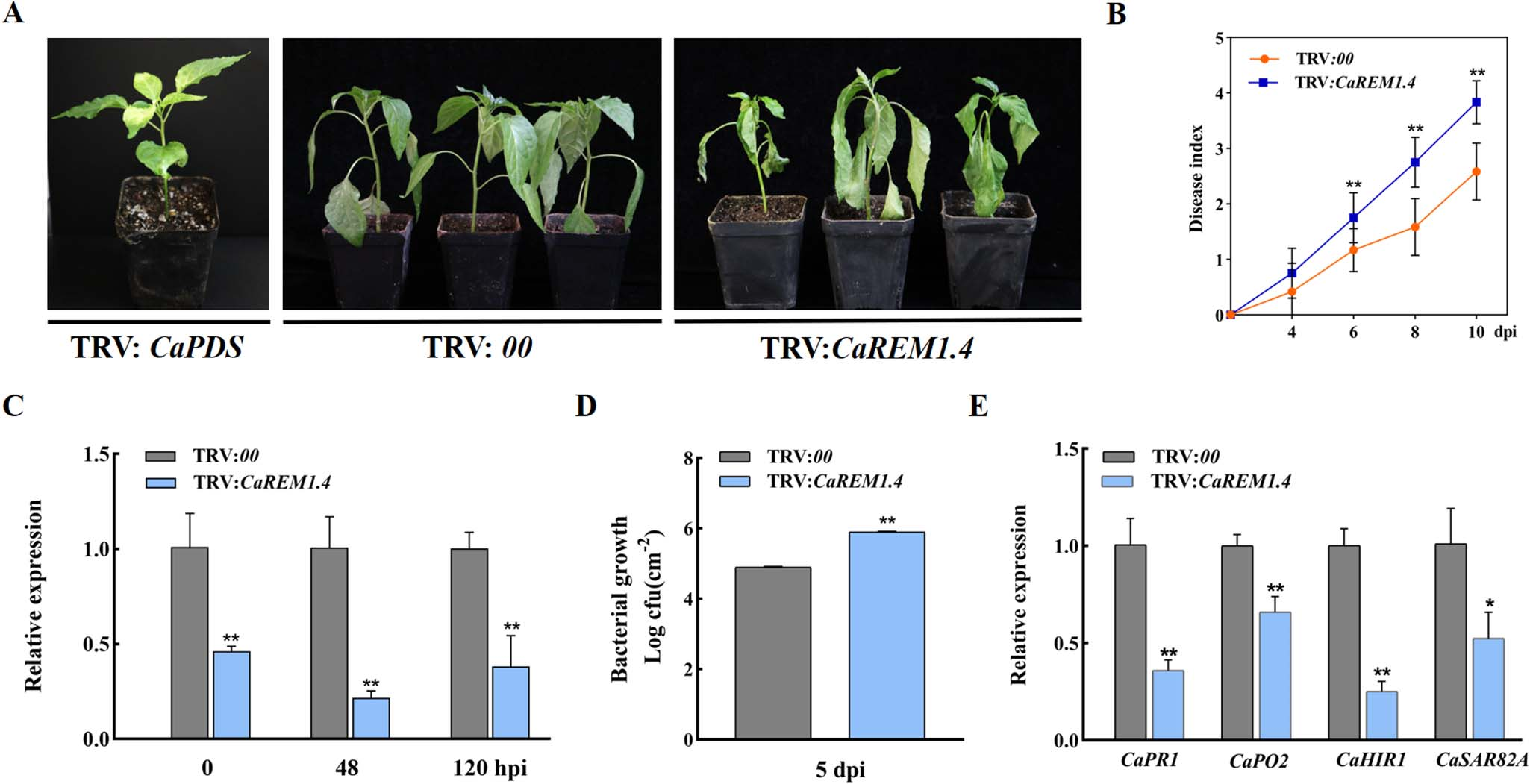
图21 VIGS实验 (Zhang et al., 2023)。
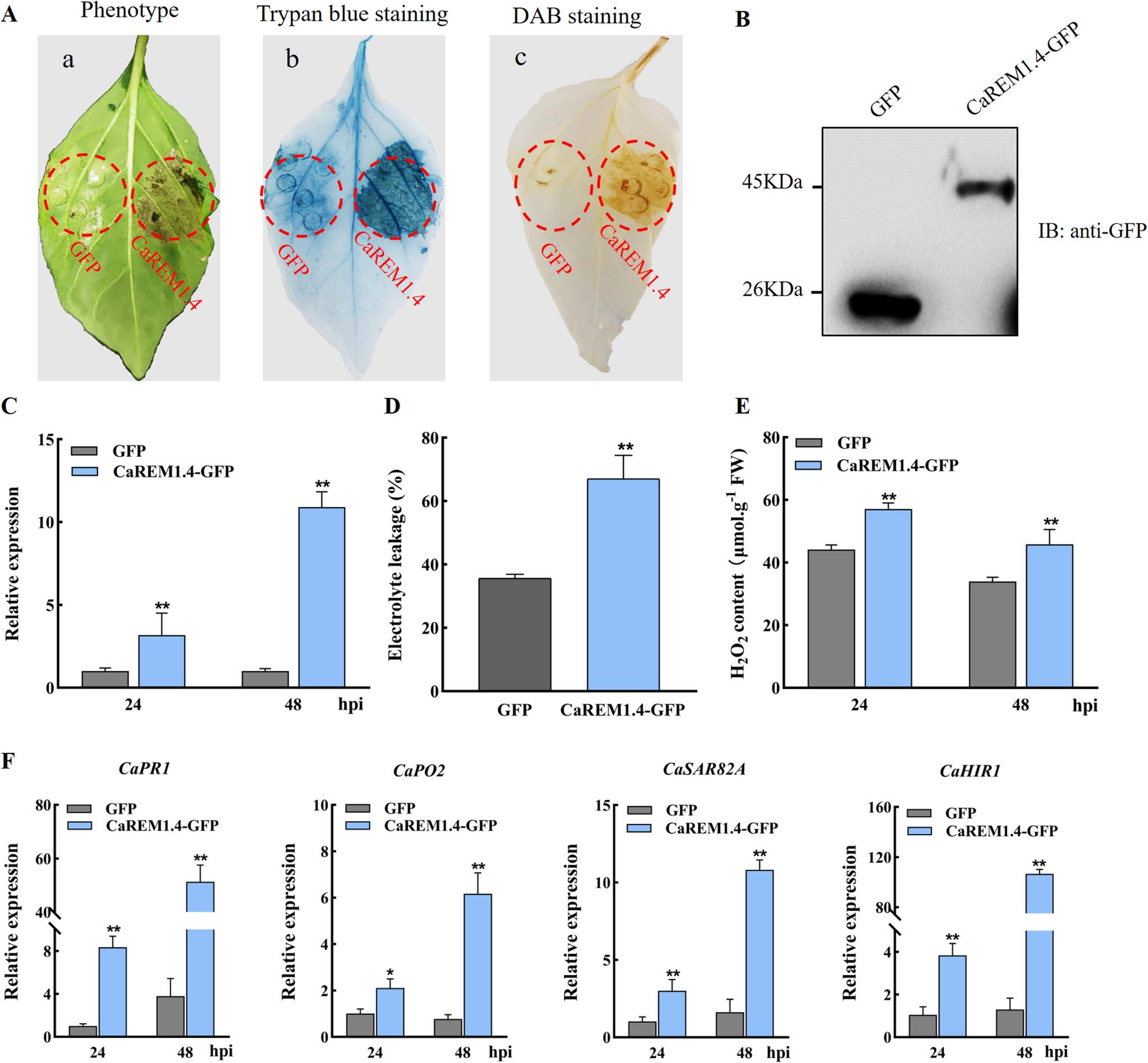
图22 瞬时过表达实验 (Zhang et al., 2023)。
在其他物种中已经证明REM蛋白与RIN蛋白互作,因此作者通过数据库的比对、搜索鉴定出辣椒中13个CaRIN4基因,再通过Split-LUC进行点对点的互作验证(见原文补充数据中),并最终选择CaRIN4-12进行后续实验验证,通过Y2H、Split-LUC、BiFC、Co-IP证明了CaREM1.4和CaRIN4-12存在相互作用(图23)。

图23 CaREM1.4和CaRIN4-12存在相互作用 (Zhang et al., 2023)。
通过在辣椒叶片中共表达CaREM1.4和CaRIN4-12(图24)以及沉默CaRIN4-12(图25)发现,CaRIN4-12能显著减少CaREM1.4产生的ROS和细胞死亡,CaRIN4-12负向调节CaREM1.4的表达,其表达量下降时可增强辣椒对青枯菌的抗性。
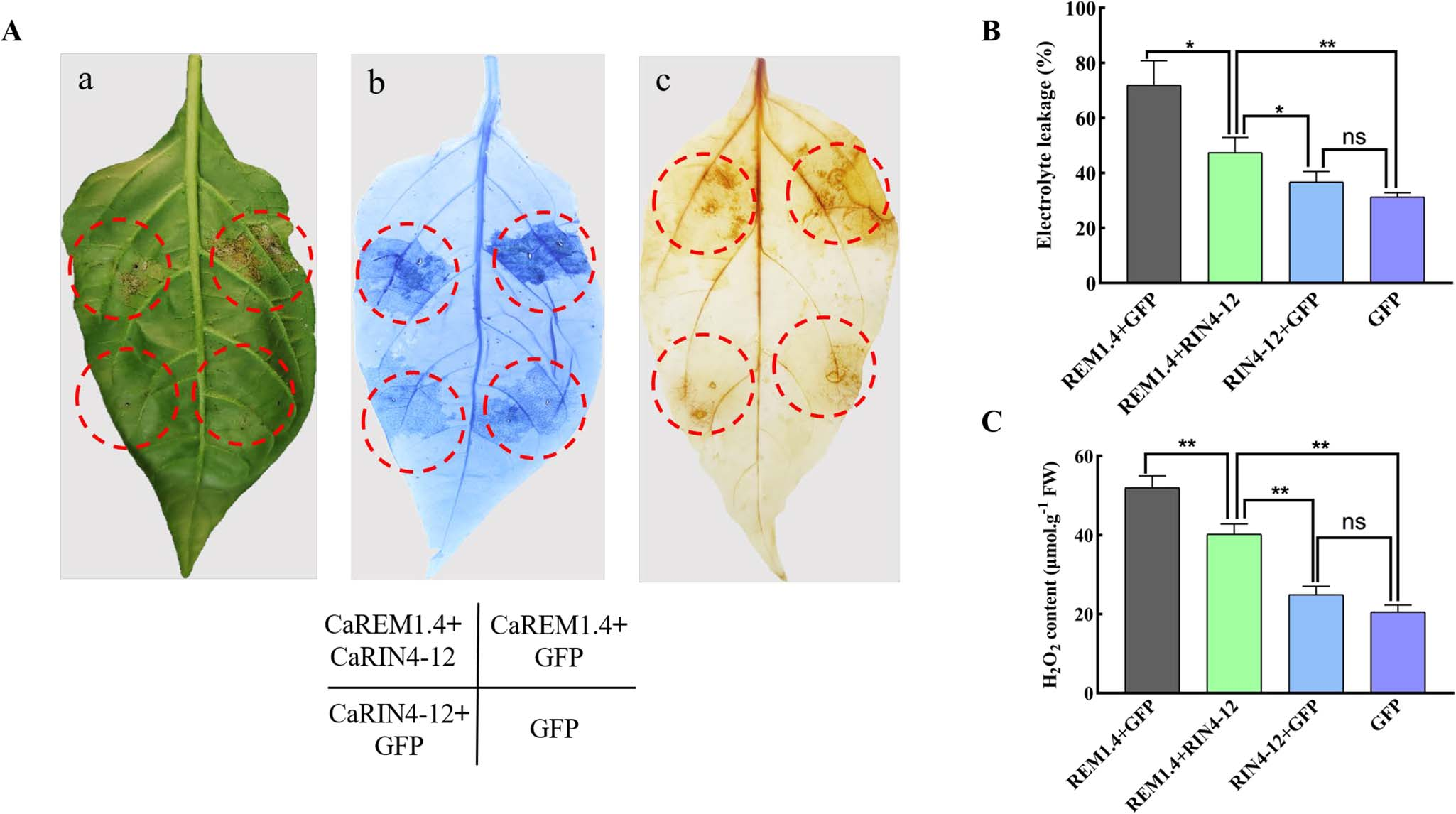
图24 共表达CaREM1.4和CaRIN4-12可减少ROS含量和细胞死亡 (Zhang et al., 2023)。
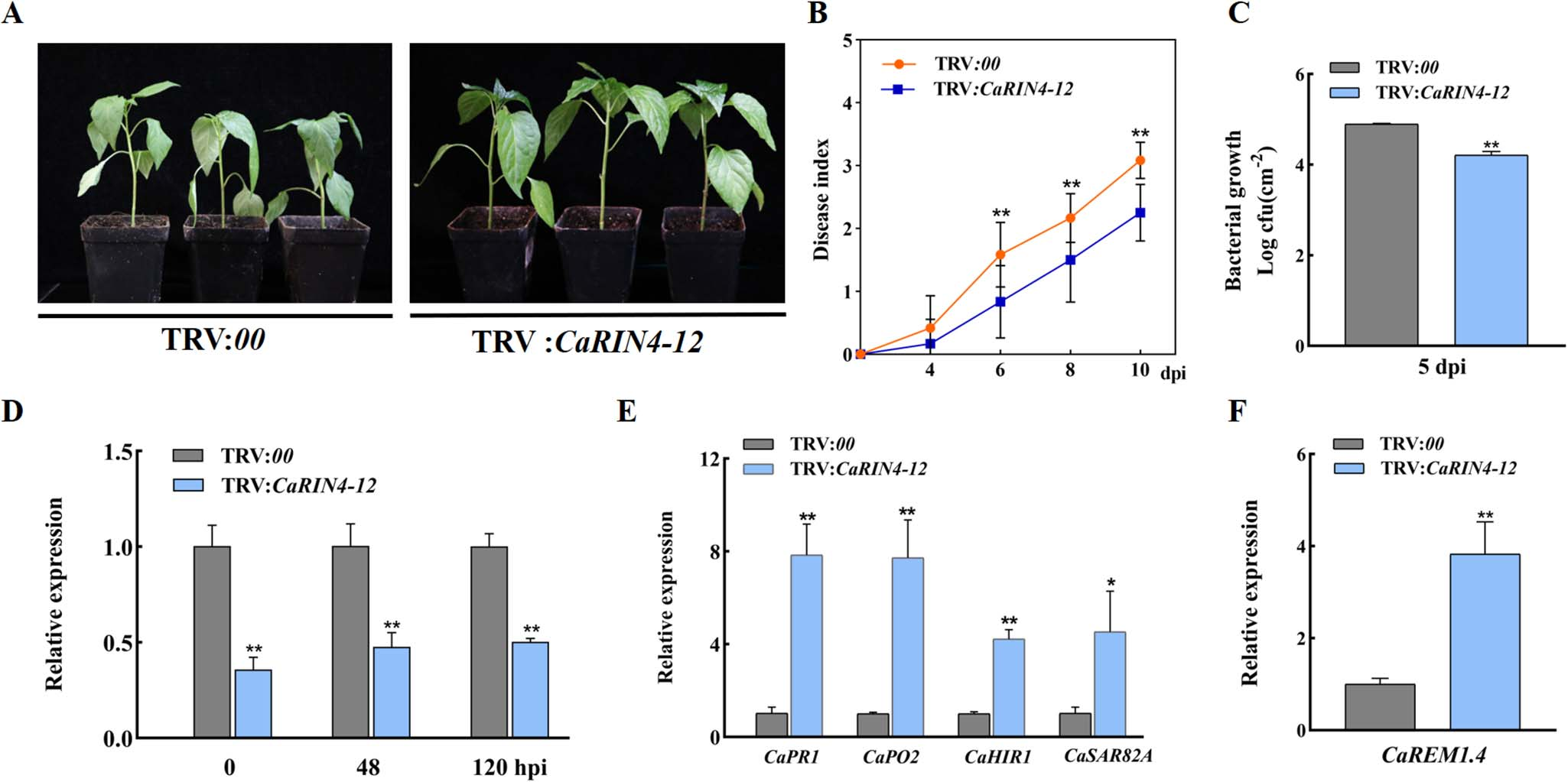
图25 沉默CaRIN4-12会降低辣椒对青枯菌的敏感性 (Zhang et al., 2023)。
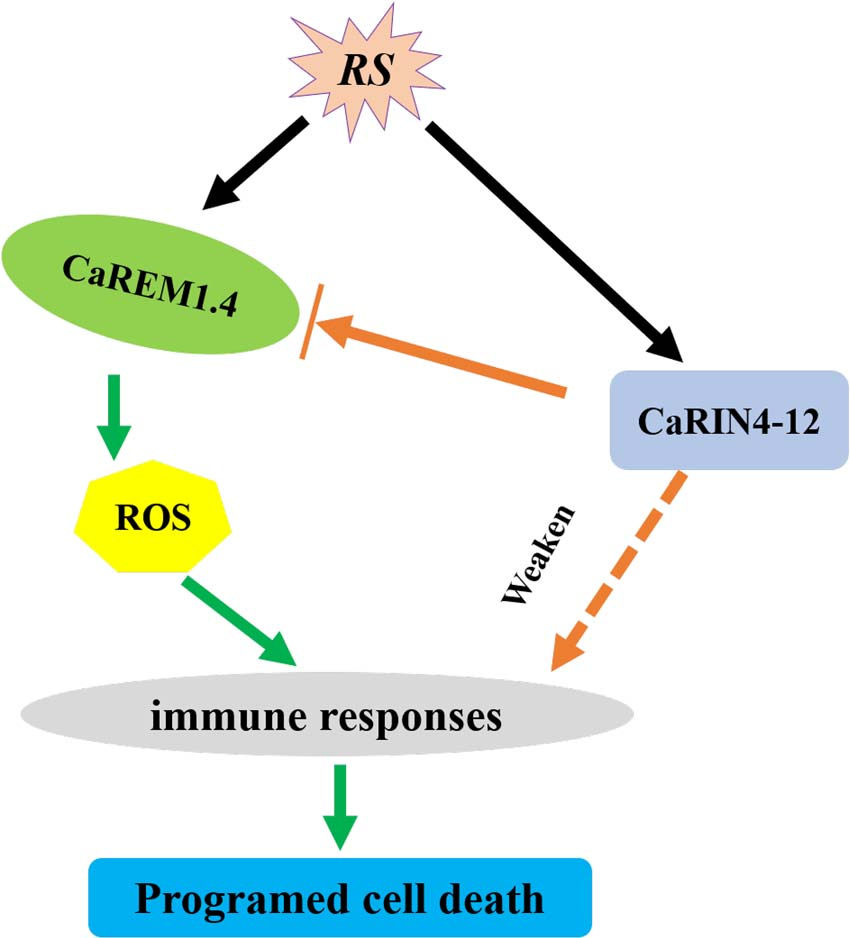
图26 CaREM1.4和CaRIN4-12介导的辣椒抗青枯病菌时的免疫反应模型 (Zhang et al., 2023)。当辣椒被青枯菌感染后,CaREM1.4通过刺激ROS产生引起细胞死亡进而诱导免疫反应,而CaRIN4-12作为植物免疫的负调控因子,可以与CaREM1.4相互作用,减少ROS的产生和细胞死亡。
伯小远将以上研究思路整理如下:
这次我们就不再放图了,只用文字来理一理作者的思路。
作者通过拟南芥WRKY33的序列在不结球白菜中寻找到了其三个同源基因BcWRKY33A、BcWRKY33B和BcWRKY33C,分析它们的功能结构域,分析WRKY33蛋白的系统发育,再通过盐胁迫处理不结球白菜“苏州青”发现BcWRKY33A受诱导程度最高,选择其用于后续研究。
通过对“苏州青”进行盐胁迫处理,发现BcWRKY33A在根中受到的诱导比叶片中高;对拟南芥稳定转化株系proBcWRKY33A::GUS染色发现BcWRKY33A主要在根中而不是叶中表达,对其进行盐处理和冷处理,发现GUS在盐处理后被诱导表达的更快、更明显。以上,表明BcWRKY33A在盐胁迫下主要在根部被诱导并发挥作用。
接着,作者通过亚细胞定位、酵母转录激活实验、双荧光素酶报告基因实验证明BcWRKY33A是转录因子。
对拟南芥稳定转化株系35S::BcWRKY33A表型进行观察,发现其比WT在盐胁迫下拥有更多的侧根、更长的根长和更高的成活率。拟南芥SALK突变体wrky33-1在盐胁迫下比WT表现出更少的侧根。在苏州青中进行VIGS实验,发现沉默BcWRKY33A的叶片具有更高的盐敏感性、盐胁迫处理后相比于对照光合能力受损更严重、丙二醛(MDA)含量、电解质渗透率、活性氧(ROS)含量水平更高。
使用酵母双杂筛库技术筛到BcHSFA4A与BcWRKY33A(包含两个保守的WRKY结构域)互作,并用BiFC进行点对点的互作验证。
接下来,对BcHSFA4A基因进行基因功能研究:检测其组织表达模式;对拟南芥稳定转化株系35S::BcHSFA4A、突变体hsfa4a-1进行表型观察;利用酵母转录激活实验、双荧光素酶报告基因实验发现BcHSFA4A具有较弱的转录激活;根据前人文献所述,猜测BcHSFA4A可能可以结合BcZAT12和BcHSP17.6A基因的启动子,通过EMSA实验证实了这个猜测。
通过EMSA、双荧光素酶报告基因实验证明BcHSFA4A结合BcZAT12和BcHSP17.6A基因启动子的能力不仅受到BcWRKY33A的影响,甚至BcHSFA4A与BcWRKY33A的相互作用能增强BcHSFA4A对下游基因表达的激活能力。在拟南芥35S::BcHSFA4A/35S::BcWRKY33A品系中发现这两个基因的共表达可以促进耐盐相关基因的高表达。
伯小远将以上研究思路整理如下:
这些例子,无一例外,都是通过反向遗传学找到的目的基因。其中,最受欢迎的候选基因是转录因子,比如案例一和案例四中的NAC、WRKY基因。当然,也有像案例三中的非转录因子基因REM,也会有少数文献比如案例二去研究lncRNA等非编码基因。我们发现,绝大多数情况下,这些目的基因在其他模式物种中有比较深入的分子机制知识积累,比如目的蛋白的互作蛋白有哪些、目的蛋白能结合哪些下游基因的启动子、某种胁迫又会诱导哪些基因发生变化等,这些研究的会比较透彻,能给作者研究的无转化体系的物种提供一定的参考。
四个案例中的物种,有些是压根没有稳定转化的体系,有些是稳定转化的体系基因型依赖太强,只限几个品种可以做稳定转化,所以作者基本都选择瞬时过表达和VIGS实验来研究基因功能,当然,有的也选择在成熟的模式植物中来辅助验证基因功能。我们也会发现,这些物种的基因数据库资源是充足的,否则很难通过反向遗传学找到候选基因以及继续向下做基因网络解析实验。所以没有成熟的转化体系,看起来并不能成为无法做分子实验、不能发好文章的理由喔!
Geng L, Yu S, Zhang Y, et al. Transcription factor RcNAC091 enhances rose drought tolerance through the abscisic acid-dependent pathway [published online ahead of print]. 2023 Jun 26. Plant Physiol. 2023;kiad366.
Wang H, Li Z, Ren H, et al. Regulatory interaction of BcWRKY33A and BcHSFA4A promotes salt tolerance in non-heading Chinese cabbage [Brassica campestris (syn. Brassica rapa) ssp. chinensis]. Hortic Res. 2022; 9: uhac113. Published 2022 May 17. doi:10.1093/hr/uhac113
Zhang Y, Guo S, Zhang F, et al. CaREM1.4 interacts with CaRIN4 to regulate Ralstonia solanacearum tolerance by triggering cell death in pepper. Hortic Res. 2023; 10(5): uhad053. Published 2023 Mar 28. doi:10.1093/hr/uhad053
Zhou R, Dong Y, Wang C, et al. LncRNA109897-JrCCR4-JrTLP1b forms a positive feedback loop to regulate walnut resistance against anthracnose caused by Colletotrichum gloeosporioides[J]. Hortic Res, 2023, 10(6): uhad086.
