病毒,尤其是植物病毒,基因组很小,编码的蛋白质非常有限,但病毒利用这些为数不多的工具完成了包括复制、转运、在宿主之间传播等功能。病毒入侵时与宿主因子相互作用,这些宿主因子可能具有抗病毒或抑制病毒的功能。宿主抑制病毒因子通常被称为宿主易感因子,在不存在宿主(或存在对病毒不起作用的亚型)的情况下宿主不敏感或不完全易感,易感性的丧失在表型上等同于抗性。多年来,研究人员利用隐性抗性基因来培育抗病毒的栽培品种,这些隐性病毒抗性基因符合编码易感因子的假说。番茄花叶病毒(PepMV)是一种单链正义RNA病毒,在全球番茄作物中流行造成了非常严重的经济损失。鉴于PepMV对经济的影响,前期已经在茄属植物中进行了抗性筛选,但成效有限。2023年7月13日,国际著名杂志Plant Biotechnology Journal杂志在线发表了题为“The tomato calcium-permeable channel 4.1 (SlOSCA4.1) is a susceptibility factor for pepino mosaic virus”的研究论文,文章中通过筛选了一系列番茄突变体鉴定到与PepMV易感性降低相关的关键突变,并成功定位到一个新的易感因子SlOSCA4.1,这一研究不仅有助于培育抗PepMV的番茄品种,并有助于我们更好地理解病毒生物学和病毒-宿主分子相互作用。

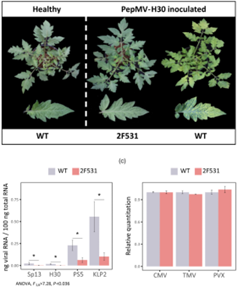
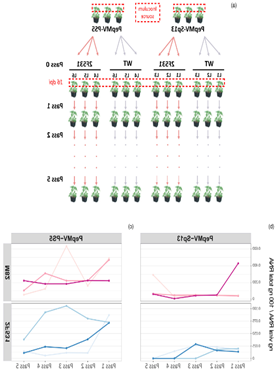
研究人员通过对来自1000 M2家族的25000棵番茄突变体进行了PepMV病毒分离株的接种和表型筛选,无症状或症状轻微的植株进行自交产生的M3家系再次接种病毒,结果发现后代家系2F531显示出100%的无症状植株,这表明该家系存在导致对PepMV失去敏感性的同源突变。PepMV的四种分离株接种2F531均未出现病症,但接种CMV,TMV,PVX等其他病毒时2F531出现了感染症状。PepMV-SP13和PepMV-PS5的两种病株在WT或2F531植物连续传代5次后,结果发现两种病毒载量随传代的波动很明显,2F531中的病毒载量总是比野生型小(在大多数情况下几乎是一个数量级)。这些结果表明家系2F531表现出的针对PepMV的抗性是具有特异性和持久性的。
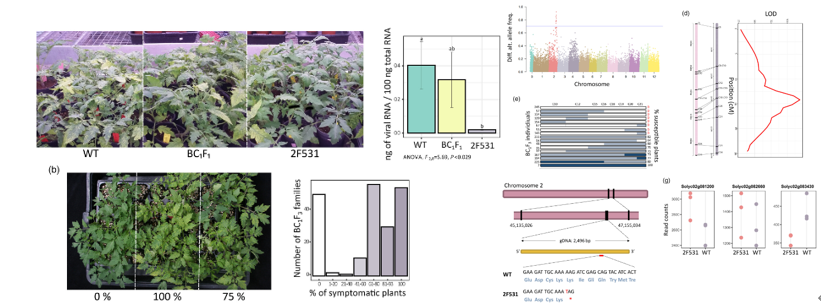
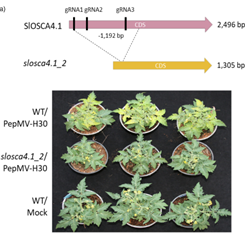
家系2F531回交得到的BC1F1植株表现出与WT植株相似的易感性表型,BC1F3的植株中所有个体要么抗性要么易感,这表明2F531的抗性是由单基因隐性遗传。作者利用BSA-HTS分析和RNA-Seq数据最终定位到了对PepMV失去敏感性的同源突变-由Solyc02g083430编码SlOSCA4.1基因。CRISPR / Cas9编辑SlOSCA4.1基因后,与WT植物相比,突变体中的PepMV病毒量积累显着减少进一步证实SlOSCA4.1是赋予PepMV易感性的基因。分析发现突变体554F2中氨基酸531处的过早引入终止密码子可能导致大部分RSN1_7TM跨膜结构域的丢失,这可能导致蛋白质功能的全部或部分丧失从而导致2F531突变表型。同样,在其N末端缺失几乎一半的蛋白后也降低了突变体对PepMV的易感性。
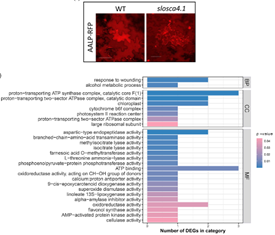
SlOSCA4.1在拟南芥直向同源物编码为AtOSCA4.1,它属于机械激活离子通道OSCA/TMEM63家族。作者提出SlOSCA4.1似乎以细胞自主的方式对PepMV起作用,因为融合蛋白mRFP-SlOSCA4.1在PepMV感染后在细胞内重新定位,在健康的本氏烟草表皮细胞中,mRFP-SlOSCA4.1 的分布模式表明为ER膜定位,而在感染后,mRFP-SlOSCA4.1重新定位为细胞内的病毒聚集体,这很可能对应于病毒复制复合物。RNA-Seq分析表明SlOSCA4.1参与了胁迫反应,可能是通过其跨膜Ca2+转运功能。SlOSCA4.1的缺失可能会阻碍Ca2+平衡的正确调节,而Ca2+平衡是PepMV最佳复制所特别需要的。SlOSCA4.1在感染期间的重新定位为可能的PepMV复制复合物与这一假设完全兼容,有待进一步测试。
文章来源:植物生物技术Pbj
