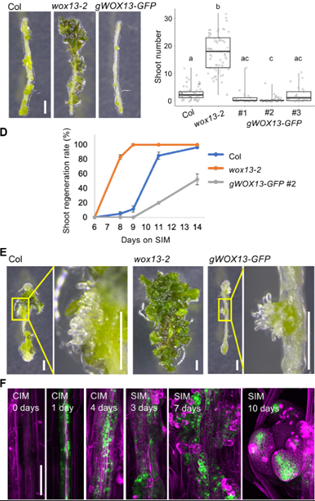
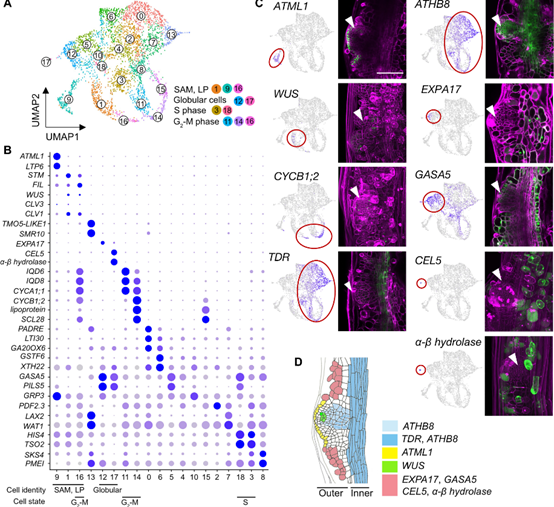
植物具有突出的再生能力,可以通过体细胞培养重建植株个体,其中的关键步骤就是茎尖分生组织(SAM:shoot apical meristem)的重头从建。目前研究人员已经对拟南芥器官再生过程中SAM启动的基因调控机制进行了广泛分子遗传学研究:在广泛使用的两步组织培养体系中,外植体首先在富含生长素的愈伤诱导培养基(CIM)上培养产生愈伤组织,然后转移到富含细胞分裂素的茎尖诱导培养基(SIM),直接上调WUS,诱导SAM 的重头从建。WUS是SAM命运获得的早期标志基因,只在一小部分愈伤组织细胞中被激活,且只有一小部分表达WUS的细胞簇最终成为SAM。同时植物也有限制愈伤组织再生的负调控,一些表观遗传调控维持靶基因位点的抑制状态,从而阻碍细胞命运从愈伤组织向SAM转变。近日,日本奈良科学技术大学科学技术研究生院生物科学部MOMOKO IKEUCHI在国际知名期刊Science Advances上在线发表了题为“WUSCHEL-RELATED HOMEOBOX 13 suppresses de novo shoot regeneration via cell fate control of pluripotent callus”的研究论文,基于Quartz-Seq2单细胞转录组揭示了WOX13在愈伤组织细胞群体的细胞身份决定中起关键作用。首先作者观察了WOX13在控制茎分生组织中的潜在作用,发现在SIM培养基中培养11天后,wox13突变体产生更多的分生组织,而将其基因组片段回补到突变体中,产生了更少的分生组织。进一步对其时空表达模式进行分析发现: WOX13在完整的黄化下胚轴中不表达,在CIM培养基中培养1天后开始表达,但转移至SIM培养基中培养10天后,WOX13在已经形成的SAM 中不表达。这些证据证明WOX13是茎再生的负调节因子,其表达在培养后诱导,并在SAM中局部减少。为了进一步探究WOX13负调控SAM形成的分子机制,作者选取在SIM培养基上培养7天正在形成SAM的材料经过细胞分选后进行单细胞转录组测序,发现了19个细胞类群,并进行了标志基因的标记,发现基因表达谱分为愈伤组织的内层和外层,外层的细胞进一步分为SAM和非分生组织细胞。进一步分析发现:WT和wox13突变体之间的差异表达至少部分归因于愈伤组织细胞组成的差异,WOX13促进了愈伤组织中非分生细胞的表达,作者推测WOX13和WUS可能是愈伤组织细胞群体中二元细胞命运的决定因素。然后作者发现WOX13和WUS间存在相互排斥的空间表达模式,提出了WOX13和WUS相互抑制的假设并加以验证。总之,作者发现WOX13与WUS构成了一个相互抑制的调控回路,在空间上分离了愈伤组织细胞群体中的微区域,这对再生效率有重要影响。文章来源:植物生物技术Pbj