在大多数生态系统中,光合作用通过将环境中的CO2转化为有机碳来形成食物链的基础。这些反应的核心是核酮糖-1,5二磷酸羧基酶/加氧酶(Rubisco),它将CO2同化为用于产生生物质的糖前体。尽管Rubisco在光合作用中起着关键作用,但它有两个局限性:1)它对中心碳代谢中的一种酶的催化速度很慢;2)它还可以催化氧合作用,这是一种使用O2而不是CO2的浪费反应。Rubisco催化速率和CO2与O2的特异性之间的权衡似乎阻止了Rubisc既快速又特异的进化。为了将氧化作用保持在可容忍的低水平,许多植物使用一种特定但缓慢的Rubisco形式,并通过产生大量的酶来弥补其缓慢的催化速率。这一策略需要大量的细胞资源,包括高达25%的叶片总氮。
一些光合作用生物通过使用CO2浓缩机制(CCM)将浓缩的CO2输送到酶中,从而克服了Rubisco的局限性。这种浓缩的CO2增加了Rubisco的周转速度,较高的CO2与O2的比率有利于羧化并抑制氧化。目前人们对了解CCM如何工作很感兴趣,这既是因为它们的重要生态作用,也因为将CCM转入到作物中具有增加产量的潜力。
根据用于浓缩CO2的中间分子的性质,CCM被分为两大类:生化和生物物理。生化CCM包括C4、C2和景天酸代谢,将CO2瞬间固定在草酰乙酸酯和苹果酸等中间有机分子中,浓缩的CO2在Rubisco附近释放出来。相比之下,在生物物理CCM中,唯一的中间分子是碳酸氢盐。生化CCM主要在植物中发现,通常涉及多细胞结构,而生物物理CCM主要在微生物中发现,并在单细胞水平上运行。
原核生物和真核生物之间的生物物理 CCM 不同。两者都依赖于一种亚细胞结构,其基质含有高浓度的 Rubisco,HCO3-中释放出浓缩的 CO2。然而,称为淀粉核(直径 1–2 µm)的真核细胞室比含有 Rubisco 的细菌细胞室,即羧基体(直径约 200 nm)大得多。此外,两种结构中的 CO2输送到 Rubisco 被认为是基于不同的原理实现的:在淀粉核中,CO2输送由类囊体膜介导。而在羧基体中,CO2产生来自HCO3-直接扩散到羧基体基质中。这篇综述重点关注淀粉核。
淀粉核是一种相分离的细胞器,在大多数真核藻类和陆地植物角苔谱系中增强光合碳同化。淀粉核介导了大约三分之一的全球 CO2固定,并且预计将淀粉核工程化到 C3作物中会促进 CO2吸收并提高产量。淀粉核通过向固定CO2的酶Rubisco提供浓缩的CO2来增强它的活性。所有的淀粉核都有一个致密的Rubisco基质,与光合作用类囊体膜相关,能提供浓缩的CO2。许多淀粉核也被多糖结构所包围,这可能会减缓CO2的泄漏。系统发育分析和形态多样性支持淀粉核趋同进化起源。大多数对淀粉核的分子认识来自于模式绿藻衣藻(Chlamydomonas reinhardtii)。衣藻淀粉核在细胞周期和响应环境诱因下表现出多种类液体行为,包括内部混合、分裂分裂、溶解和凝结。淀粉核的组装和功能是由CO2可获得性和光照诱导的,尽管已经确定了转录调控因子,但翻译后调控仍有待研究。
相关内容以“The pyrenoid: the eukaryotic CO2-concentrating organelle”为题于2023年6月4日在国际著名期刊《The Plant Cell》在线发表。本文对衣藻中淀粉核的功能、结构、组成和动态调控进行了综述,并对其他物种的淀粉核进行了推断。
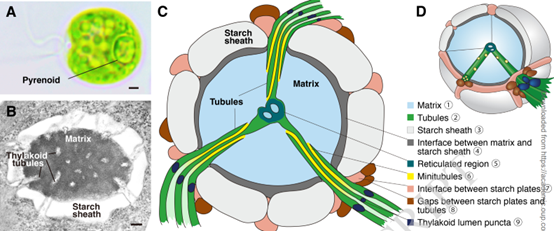
图1.衣藻淀粉核的结构。(A) 衣藻淀粉核在光学显微镜下可见。比例尺:1 μm (B) 衣藻蛋白核由三个主要隔间组成:Rubisco 基质、类囊体小管和淀粉鞘。比例尺:200 纳米。(C) 二维和 (D) 核蛋白的三维模型,显示主要隔间和蛋白质外围结构。
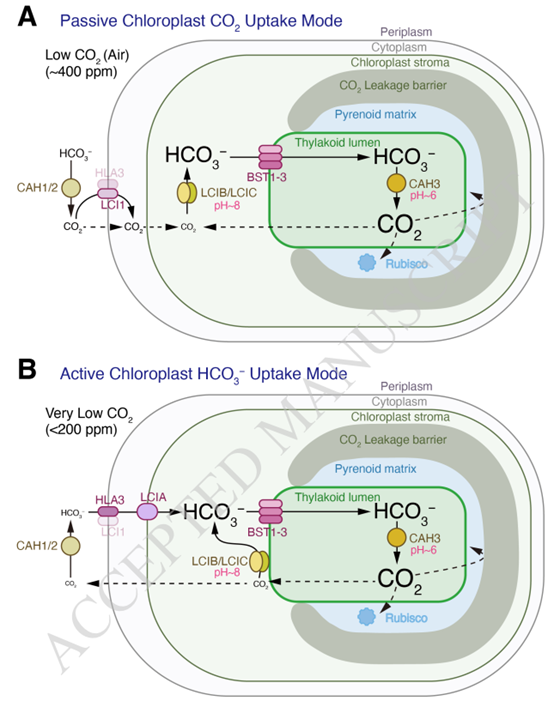
图2. 基于核蛋白的 CO2 浓缩机制 (CCM) 的工作原理。
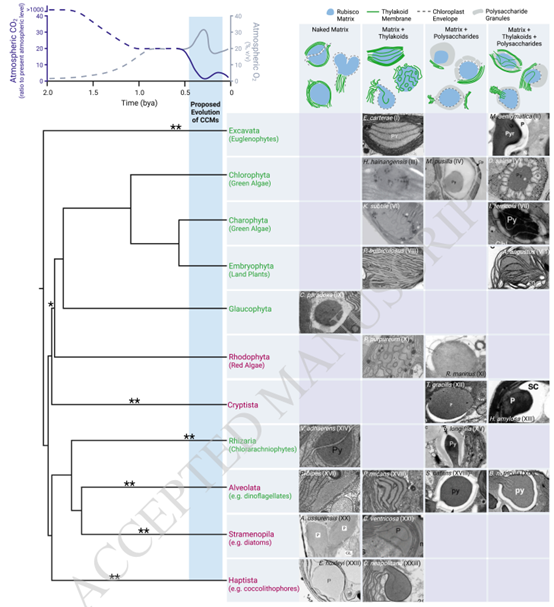
图 3. 淀粉核似乎已随着大气中二氧化碳浓度的下降而趋同进化。
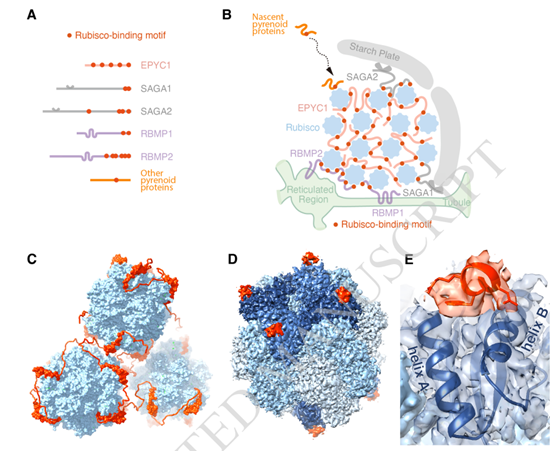
图4.一个常见的Rubisco结合基序介导了淀粉核的主要隔室的组装。