Metacaspases(MCAs)与caspases和paracaspases都是是半胱氨酸蛋白酶家族的一部分。MCAs在整个植物、真菌、原生生物和细菌中都是保守的,而caspases和paracaspases则存在于哺乳动物中。MCAs根据它们的域组织进行分类:I 型有一个 N端结构域,而 II 型没有,但是后者有一个更长的接头连接 p20 和 p10 区域。III 型MCAs最近在浮游原生植物的基因组中被发现,并且在 p20 之前具有 p10 区域的重排,在其他方面具有与 I 型和 II 型相似的活性特征。保守的 p20 和 p10 区域共同构成核心蛋白酶结构域。

大多数 MCAs 需要钙离子 (Ca 2+ ) 和中性 pH 值才能发挥活性,II 型MCAs 的一个亚组明显例外,它仅存在于维管植物中,它们在弱酸性 pH 值(约 pH 5.5)下发挥最佳功能并且不需要 Ca 2+。它们除了在病原体诱导的程序性细胞死亡(PCD)中的作用,也在细胞衰老和蛋白质聚集物清除、细胞清除、伤口诱导的损伤相关分子模式信号和发育性细胞死亡事件中发挥作用。
近日,PNAS杂志在线发表了根特大学植物生物技术与生物信息系Simon Stael团队题为“Structure function study of a Ca2+-independent metacaspase involved in lateral root emergence”的研究论文, 他们报告了拟南芥中一类钙离子非依赖性MCA—— AtMCA-IIf 的结构,并为一类含 TDP 的小分子对MCAs的抑制作用提供了基本原理。
迄今为止,I 型MCA的两种蛋白质晶体结构已从酿酒酵母和布氏锥虫(TbMCA-Ib)中发表,而一种来自拟南芥II 型钙依赖性 MCA (AtMCA-IIa)也被发表. 所有三个结构的核心是一个典型的半胱天冬酶/血红蛋白酶 alpha-beta-alpha 夹心折叠。在 I 型MCA结构中,蛋白质的 N 端部分结合在活性位点,这解释了为什么这些酶在去除 N 端前结构域之前没有完全激活。此外,构成催化二元体的半胱氨酸和组氨酸残基相距太远,并且处于催化组氨酸不能作为负责半胱氨酸去质子化的碱基的构象,否则通常被认为是其他半胱氨酸蛋白酶如木瓜蛋白酶的机制。这个难题本身也存在于催化机制尚未完全解释的半胱天冬酶中。在 AtMCA-IIa 结构中,部分连接子区域结合在活性位点,这解释了该区域需要自动切割以进行激活。AtMCA-IIa 的分解结构不含 Ca 2+,因此尚不清楚 Ca 2+在 II 型 MCA 中的结合位置,以及它如何触发激活。与 I 型 MCA 中的难题类似,AtMCA-IIa 结构也显示酶的非活性形式,催化残基相距太远。此外,对于Ca 2+非依赖性MCA,缺乏代表性的晶体结构,这进一步阻碍了对MCA激活机制的理解。
侧根构成植物根系的重要组成部分,确保植物最佳的养分吸收、水分吸收和土壤固定。侧根生长受植物激素生长素的严格调控,在生长素的控制下,这些初始细胞发育成侧根原基 (LRP),然后通过覆盖的内皮层和皮层生长,直到最终突破表皮并从初生根中长出。. 在拟南芥中,AtMCA-IIf在起始 LRP 前的内皮层中的特异性时空转录诱导表明其参与内皮层的 PCD 以清除原基发育过程。然而,很可能是由于功能冗余,单个AtMCA-IIf/突变体在侧根出现期间不会遇到问题。拟南芥基因组包含三个I型和六个II型MCA。
在本研究,他们采用化学生物学方法来解MCAs 在侧根形成中的作用。首先,他们解析了 AtMCA-IIf 的晶体结构,AtMCA-IIf 是拟南芥中唯一的不依赖 Ca 2+的MCA。然后,他们开发了体外化学筛选,并鉴定了一系列具有最小硫代二氢嘧啶二酮 (TDP) 结构的小分子 MCAs 抑制剂,其中一些可以特异性抑制 AtMCA-IIf 的活性。最后,他们表明含有 AtMCA-IIf 特异性 TDP 抑制剂 (TDP6) 可有效抑制体内 LRP 出现。总之,AtMCA-IIf 的结构的解析有助于更好地了解Ca 2+非依赖性MCA和相关蛋白酶(包括哺乳动物中的 caspases 和 paracaspases)的激活机制,并且抑制剂可以构成开发靶向 metacaspases 的工具和药物的基础。
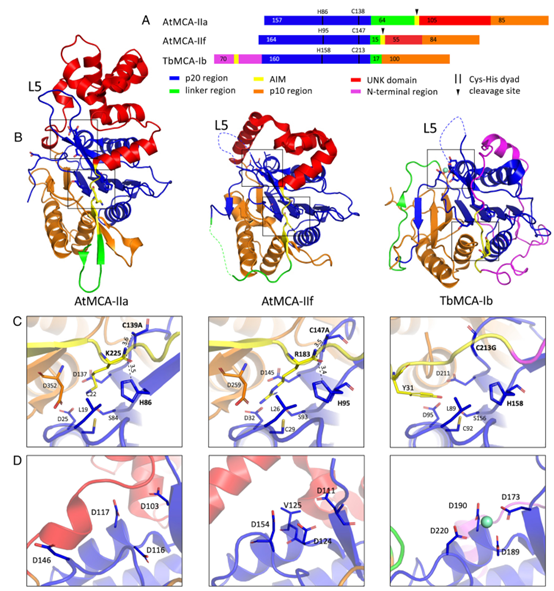
AtMCA-IIf C147A晶体结构与现有MCA结构的比较
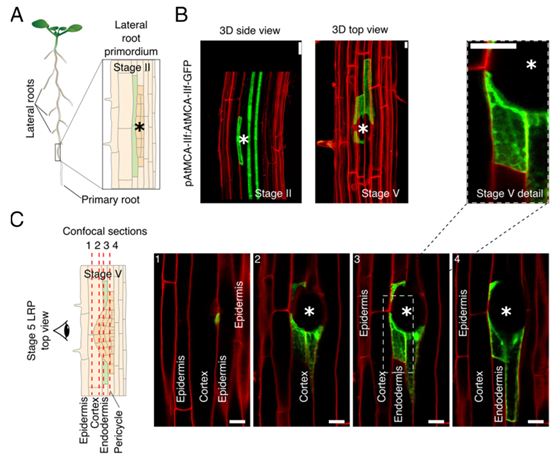
AtMCA-IIf在拟南芥LRP上的内胚层细胞中特异性表达
文章来源:植物生物技术Pbj
