对全基因组合成生物学的讨论通常忽略了重建着丝粒的问题,着丝粒是基因组中长而重复的区域,通常它发挥功能时会与CENP-A/CENH3或其他组成型着丝粒蛋白结合来确保正常的染色体分离。人工合成的着丝粒在体内发挥作用还比较困难。
2023年3月16日,Nature Plants杂志在线发表了美国佐治亚大学的一篇题为“Synthetic maize centromeres transmit chromosomes across generations”的研究论文。该文在玉米中基于对LexA系统的使用上,证明了将CENP-A/CENH3靶向合成重复序列以激活有丝分裂形成稳定的着丝粒的基本原理适用于植物界。并进一步证明,以这种方式改造的着丝粒可以成为修饰基因组的永久特征。在缺乏原始激活蛋白的情况下,新创建的玉米合成着丝粒可以在几代中驱动独立的染色体分离。
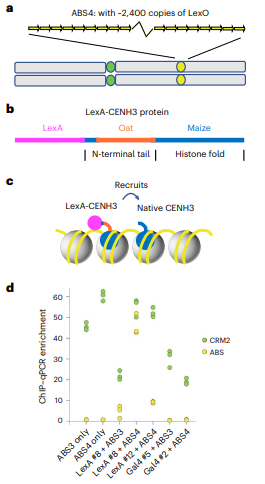
在天然的玉米着丝粒上,CENH3稀疏分布在长达2 Mb的区域。本研究中将CENH3招募到一个约377 kb的ABS区域中(图1a),并观察到CENH3扩散到连接的质粒序列和侧翼染色质上(高达~1750 kb),其中第一代(v1)结构由一个约1.7 kb的原生CENH3启动子片段组成,驱动嵌合蛋白作为无内含子的开放阅读框。将构建体转化到玉米中,与携带ABS位点的株系(3号染色体上的ABS4或ABS3)杂交,并使用玉米特异性CENH3抗体对杂合子后代进行染色质免疫沉淀(ChIP)。采用ChIP – quantitative real-time polymerase chain reaction (qPCR)和ChIP – seq测序(ChIP – seq)对结果进行分析(图1d)。LexA-CENH3(v1)显示ABS富集,而Gal4-CENH3(v1)未显示ABS富集(图1d)。这些结果表明,Lex-CENH3(v1)与ABS相互作用,并将天然CENH3招募到ABS序列中。然而,一个大的结合平台并不能保证染色体的稳定遗传,特别是在着丝粒形成后。从基因筛选的有限数据表明,经过减数分裂的含有ABS的片段中,只有约13%(3/23)会在后代中成为新染色体。
图一:LexA-CENH3(v1)将原生CENH3招募到ABS4
此外,重组的LexA-CENH3转基因植株也证明LexA-CENH3(v2)可以替代原生的CENH3,在所有组织中都能正常表达,且不干扰功能性着丝粒的组装。虽然在减数分裂中可以观察到的许多明显断裂的染色体片段可能已经丢失或退化,但更多的染色体片段可能包含在末期核中,它们可以传递给下一代。这些片段可能还带有CENH3或其他染色质标记,可以使ABS4作为新染色体的着丝粒。
图2:LexA-CENH3(v2) ABS4植物的分离情况。
来自免疫定位研究的数据表明,功能性着丝粒大小与基因组大小呈正相关。作者在大基因组植物(如玉米)中成功地设计了着丝粒,这预示着在小基因组植物(如拟南芥)中也能成功,在这些植物中,可能较小的LexO重复序列就足以形成更稳定的着丝粒。通过将新生着丝粒的形成与染色体臂的精确切割结合起来,而不是像之前描述的那样依赖于随机的染色体断裂,有可能提高新染色体形成的频率。在未来,人工合成着丝粒有可能成为解决生物经济和粮食安全挑战的重要工具。