植物对环境中大多数病原微生物都具有最常见的抗病性,这种针对非适应性病原体的植物免疫反应称为非寄主抗性 (Nonhost resistance,NHR)。解析NHR 的分子机制可以为作物抵抗病原微生物提供有效策略。有研究表明,病原体入侵前后植物的防御机制都与NHR密切相关。在与NHR相关的多层植物免疫策略中,核苷酸结合富含亮氨酸重复序列(NLR)蛋白被认为是 NHR 的重要组成部分,特别是在近代分化的植物物种中。非寄主抗性 (NHR) 是针对非适应性病原体的强大植物免疫反应。许多非适应性病原体的NLR蛋白已被鉴定,但是驱动 NHR稳健性的潜在分子机制仍然是未知的。
近日,韩国首尔国立大学农业与生命科学学院植物免疫研究中心Soohyun在Plant Biotechnology Journal上发表了一篇题为“Nucleotide-binding leucine-rich repeat network underlies nonhost resistance of pepper against the Irish potato famine pathogen Phytophthora infestans”的研究论文,该研究鉴定了致病疫霉效应子的多种遗传因素,以提高辣椒对晚疫病的非寄主抗性,为发展作物的持久抗性提供参考策略。

辣椒以高度局部化的细胞死亡响应致病疫霉的入侵,这是NLR介导的抗性的表型标志(图 1a)。为了鉴定辣椒NLRs蛋白识别的致病疫霉效应子,作者使用马铃薯病毒 X (PVX) 介导的基因表达系统测试了候选效应子是否会触发非寄主辣椒植物的超敏反应(HR)。作者筛选了57个核心 RXLR 效应子,这些效应子在四个致病疫霉分离株(T30-4、NL07434、P17777 和 06_3928A)中较为保守,并在感染马铃薯后 2-3 天内表达。将每个 RXLR 效应子的重组 PVX 病毒粒子涂抹到两种辣椒Criollo de Morelos 334 (CM334) 和 Early California Wonder (ECW) 的叶子上,发现57个效应子中有6个效应子持续诱导两种辣椒的细胞死亡(图 1b)。相比之下,尽管在 CM334 和 ECW 中检测到 GFP 荧光,但 PVX-GFP 并未引起任何特定症状。在6个效应子中,有5个是 Avrblb2 (PITG_20300) 及其旁系同源物 (PITG_04085、04090、18683、20301),这些效应子均未导致本氏烟草中的细胞死亡(图 1b)。
因此,我们还在 CM334 中使用农杆菌介导的瞬时表达筛选效应子,并鉴定出其他诱导细胞死亡的致病疫霉效应子,包括 Avr2、Avrblb1和 Avrvnt1 (图 1c )。这些结果表明,NLR蛋白与多种效应子的互作可能是辣椒抗致病疫霉的NHR的基础,这与之前的研究一致。已知候选效应子 Avr1、Avr2、Avrblb1、Avrblb2 和 Avrvnt1 具有源自野生茄属植物的相应 NLR,尽管多个致病疫霉分离株已经对相应的茄属植物 NLRs(R1、R2、Rpi-blb1、Rpi-blb2 和 Rpi-vnt1)产生抗性, 但这些分离菌株在自然条件下均不能感染辣椒。因此,我们试图鉴定出能够识别Pi效应子的辣椒NLRs,并研究辣椒NLRs如何促进对致病疫霉的强大NHR。
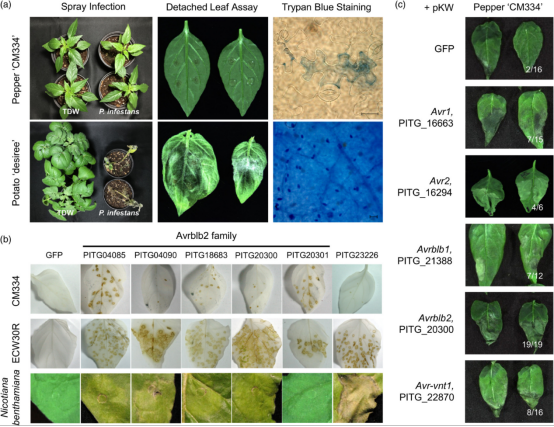
图1 致病疫霉效应子引发非寄主辣椒的HR细胞死亡
为了识别对候选效应子有反应的辣椒 NLRs,我们使用了一种反向遗传学方法,将本氏烟草作为替代系统,而不是通过将具有不同细胞死亡表型的辣椒与Pi效应子杂交来产生多个种群。这些辣椒 NLR 与本氏烟叶片上的每个候选效应子共表达(通过农杆菌渗入以 1:1 的比例表达)。特别是Avr1、Avr2、Avrblb1、Avrblb2 和 Avrvnt1 效应子已知用于相应的茄属植物 NLR。因此,与 R1 (CNL-G3)、R2 (CNL-G5)、Rpi-blb1 (CNL-G7)、Rpi-blb2 (CNL-G1)(和 Rpi-vnt1 (CNL-G11)用每个相应的效应子进行了测试(图 2a)。结果表明,辣椒具有多个识别致病疫霉效应子的 NLRs,这些 NLRs 将有助于辣椒对致病疫霉的NHR。
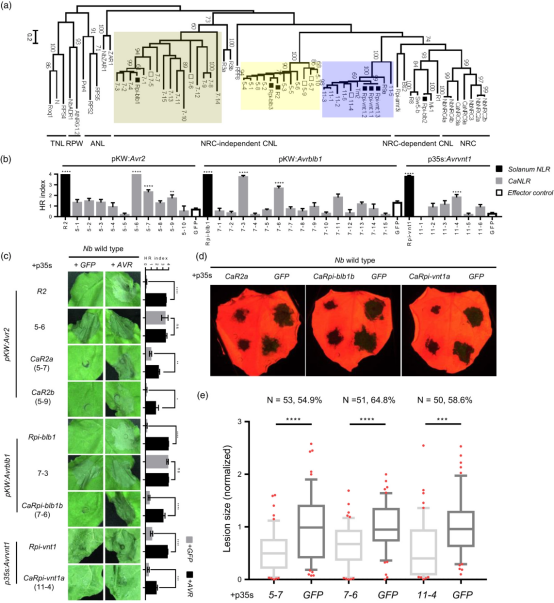 图2 本氏烟中与辣椒同源的NLRs引发HR并赋予其对致病疫霉的抗性
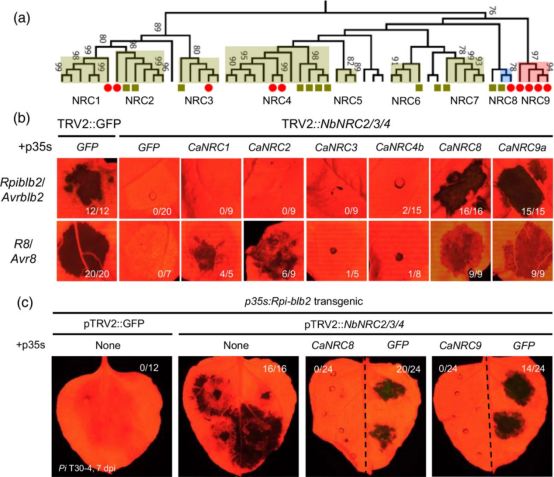
为了识别CaNRC,作者使用四种茄科植物(马铃薯、番茄、辣椒和本氏烟)的 NLR 进行了系统发育分析。作者观察到一个独特的分支,包括所有功能特征的 NRC,如 NbNRC2/3/4 和 SlNRC1,与 NRC 依赖传感器 (NRC-S) ) 可能从祖先的 NLR扩展而来(图 3a,S8)。除了辣椒的 NRC4 同源物 (Ca06g12190) 外,所有CaNRC在辣椒叶转录组数据中均以显著水平表达。NRC 进化枝显示出清晰的二分结构,分为两个由 NRC1/2/3 和 NRC4 同系物组成的子进化枝。NRC1/2/3 同系物在四种茄科植物中普遍保存。相比之下,NRC4 同源物表现出物种多样性(图 3a,S8),表明每个茄科物种在物种形成后独立进化出 NRC4。这些结果表明辣椒进化出功能相似NRC4类辅助NLR,但这些谱系特异性 CaNRC与其本氏烟草的同系物相比遵循不同的进化轨迹。这可能导致辣椒 NRC-S NLR 和NbNRC之间的功能不相容。
图3 辣椒进化出谱系特异性NRC以补充NbNRC4功能
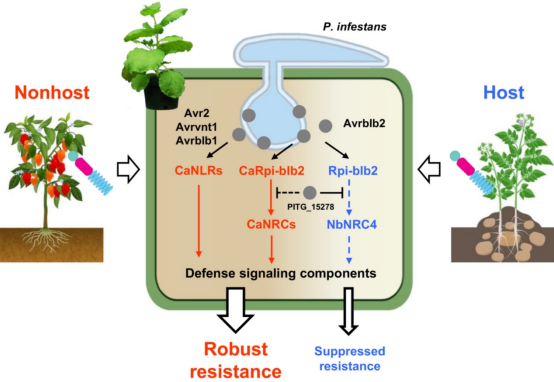
这项研究表明,可以利用非寄主植物的 NLR 网络来赋予对非适应性病原体的抗性。非寄主辣椒具有多个与茄属NLRs 同源的 NLRs ,并识别非适应性病原体致病疫霉的相同效应物(图 2、4)。因此,作者提出了一个模型,其中辣椒的 NHR 是多个 R 基因叠加的结果,其中一些基因逃避了致病疫霉效应子的抑制(图 6),病原体效应子之间保守的结构(由相似的 NLR 直接识别)或功能(与由 NLR 保护的相似宿主目标相互作用)同源性,可能使植物能够识别源自适应和非适应病原体的效应子。
图6 NLR网络介导的辣椒NHR防御致病疫霉的机理模型
综述所述,当非寄主植物(辣椒)与寄主植物(土豆)亲缘关系密切时,多个NLRs介导了对非适应病原体效应物的识别。这一结果与先前关于非寄主/寄主植物的进化距离与受体介导的非寄主抗性的分子机制之间关系的假设一致。此外,这些非寄主植物衍生的部分 NLR可能对病原体的免疫抑制更具耐受性,而致病疫霉已经对来自原始宿主(马铃薯、番茄)密切相关的野生茄属植物的NLR产生抗性。因此,进一步鉴定和解析源自远缘非寄主植物的 NLR 将为开发针对致病疫霉等破坏性病原体的持久抗性作物提供有效策略。
图2 本氏烟中与辣椒同源的NLRs引发HR并赋予其对致病疫霉的抗性
为了识别CaNRC,作者使用四种茄科植物(马铃薯、番茄、辣椒和本氏烟)的 NLR 进行了系统发育分析。作者观察到一个独特的分支,包括所有功能特征的 NRC,如 NbNRC2/3/4 和 SlNRC1,与 NRC 依赖传感器 (NRC-S) ) 可能从祖先的 NLR扩展而来(图 3a,S8)。除了辣椒的 NRC4 同源物 (Ca06g12190) 外,所有CaNRC在辣椒叶转录组数据中均以显著水平表达。NRC 进化枝显示出清晰的二分结构,分为两个由 NRC1/2/3 和 NRC4 同系物组成的子进化枝。NRC1/2/3 同系物在四种茄科植物中普遍保存。相比之下,NRC4 同源物表现出物种多样性(图 3a,S8),表明每个茄科物种在物种形成后独立进化出 NRC4。这些结果表明辣椒进化出功能相似NRC4类辅助NLR,但这些谱系特异性 CaNRC与其本氏烟草的同系物相比遵循不同的进化轨迹。这可能导致辣椒 NRC-S NLR 和NbNRC之间的功能不相容。
图3 辣椒进化出谱系特异性NRC以补充NbNRC4功能
这项研究表明,可以利用非寄主植物的 NLR 网络来赋予对非适应性病原体的抗性。非寄主辣椒具有多个与茄属NLRs 同源的 NLRs ,并识别非适应性病原体致病疫霉的相同效应物(图 2、4)。因此,作者提出了一个模型,其中辣椒的 NHR 是多个 R 基因叠加的结果,其中一些基因逃避了致病疫霉效应子的抑制(图 6),病原体效应子之间保守的结构(由相似的 NLR 直接识别)或功能(与由 NLR 保护的相似宿主目标相互作用)同源性,可能使植物能够识别源自适应和非适应病原体的效应子。
图6 NLR网络介导的辣椒NHR防御致病疫霉的机理模型
综述所述,当非寄主植物(辣椒)与寄主植物(土豆)亲缘关系密切时,多个NLRs介导了对非适应病原体效应物的识别。这一结果与先前关于非寄主/寄主植物的进化距离与受体介导的非寄主抗性的分子机制之间关系的假设一致。此外,这些非寄主植物衍生的部分 NLR可能对病原体的免疫抑制更具耐受性,而致病疫霉已经对来自原始宿主(马铃薯、番茄)密切相关的野生茄属植物的NLR产生抗性。因此,进一步鉴定和解析源自远缘非寄主植物的 NLR 将为开发针对致病疫霉等破坏性病原体的持久抗性作物提供有效策略。