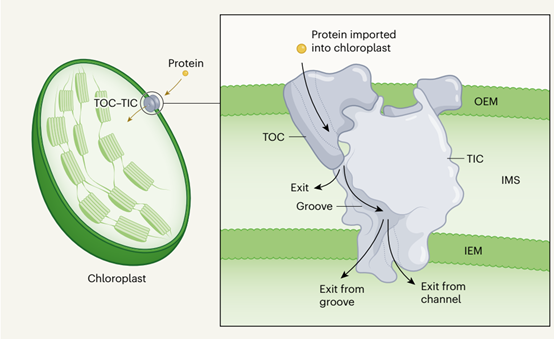
叶绿体是存在于陆生植物和藻类中的细胞器,它含有数百种负责光合作用的不同蛋白质,大多数叶绿体蛋白由核基因组编码,在细胞质中合成并含有叶绿体靶向信号。要进入叶绿体,此类蛋白质必须穿过细胞器周围的外膜和内膜(分别为 OEM 和 IEM)。OEM上的蛋白质复合物(称为TOC)和IEM上的蛋白质复合物(称为TIC)形成了复杂的机制,介导蛋白质在两层膜上的转运。以叶绿体为目标的蛋白质在TOC复合物的帮助下被识别并从细胞质中穿过OEM,TOC复合物包含两个蛋白质Toc34和Toc159,它们作为叶绿体靶向信号的表面受体,以及蛋白质Toc75,它形成一种称为β桶的结构,被认为是蛋白质输入通道。

近日,Nature杂志在线发表了日本京都产业大学生命科学学院和蛋白质动力学研究所Takashi Hirashima 和Toshiya Endo两位教授题为“Path unveiled for protein entry into chloroplasts”的研究论文, 他们利用冷冻电子显微镜测定蛋白质导入藻类叶绿体细胞器的机械结构,为了解叶绿体如何形成打开了新的篇章。
低温电镜结构的第一个惊喜是,Toc75与另一种蛋白质Toc90(也称为Toc120)组装在一起,在OEM中形成一个大型混合跨膜β桶,为蛋白质进入提供了一条途径。另一个奇特和意想不到的观察结果是,绿藻特定的OEM蛋白Ctap4(也称为Toc39)形成了第二个膜嵌入β桶,距离TOC复合体的主要部分约12埃,这个小桶的作用还有待确定。
TOC-TIC超复合物的一个显著特征是TOC和TIC复合物在IMS中形成了一个连锁结构,紧密地将两个复合物结合在一起。在植物叶绿体中,TOC和TIC复合物只有在易位蛋白的存在或桥接蛋白Tic2368的帮助下才能形成稳定的超复合物。然而,在缺乏这些蛋白质的绿藻叶绿体中形成了相对稳定的TOC-TIC超复合体。这种超复合物是稳定的,没有易位蛋白,它是否有一种桥接蛋白是未知的。低温电镜结构表明,叶绿体编码的Tic214蛋白和其他Tic蛋白共同折叠,构建了独特的IMS结构,并与TOC组分的IMS结构域相关联。此外,Tic214的一个段甚至进一步扩展,从IMS端进入TOC混合桶。然而,Tic214是如何在叶绿体内合成并被整合到复杂的IMS结构中,这还是一个谜。
TOC和TIC复合物的紧密结合将创造一个从细胞质一直到叶绿体内部的连续的蛋白质运输路线。这种途径可以防止高度疏水膜蛋白在IMS中形成错误折叠的聚集体。然而,将一些可溶性蛋白质运输到IMS也可能是一个挑战。为了克服这一问题,可溶性的IMS蛋白可能会从TOC混合桶的侧边开口进入IMS,而靶向叶绿体内部的蛋白质可能会沿着IMS结构的表面凹槽,然后到达TIC复合物膜嵌入部分的蛋白质运输路线。与TOC复合物相反,TIC复合物的膜嵌入部分由Tic20、Tic214和其他TIC蛋白提供的约15个TM螺旋组成。来自Tim20和Tim214的TM螺旋在TIC复合物的膜嵌入部分共同形成狭窄的导管。蛋白质可以潜在地通过这个通道,如果它经历一个构象变化,以扩大在收缩的位置。或者,蛋白质可能沿着面向IEM的TIC复合物的表面槽移动。
结合质谱分析,TOC-TIC超复合物的低温电镜结构揭示了以前未知的主要针对绿藻的成分。然而,低温电镜结构中缺少一些潜在的成分,而且结构中还包含一些尚未被识别的片段。因此,更深入地了解各组分的精确功能,从陆地植物中获得TOC-TIC系统的高分辨率结构,以及TIC相关的运动复合体,将为针对叶绿体形成的理解增添更多篇章。