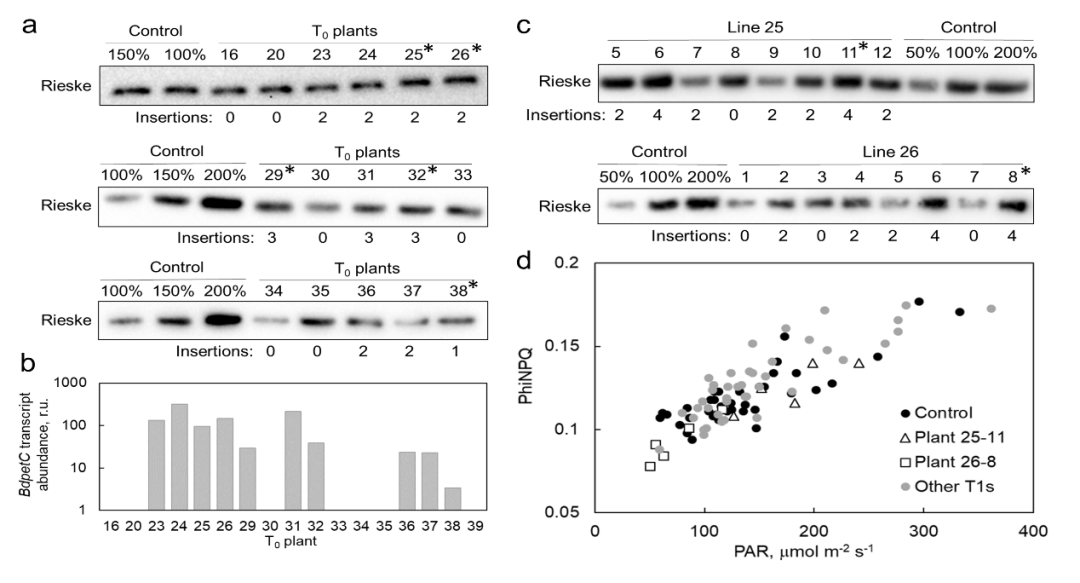
与野生型相比,转基因T0株系25、26、29和32显示出相对较高的Rieske丰度。25和26株系的T1纯合子植株与对照植株相比,具有更高的Rieske叶含量。此外,纯合子植株25-11和26-11在环境光下检测时,非光化学猝灭(NPQ)表现出相对较低的水平,这与S. viridis过表达Rieske中报道的NPQ表型一致。25-11、26-11和32-14的子代,被用于进一步的实验,以下称为转基因系25、26和32。
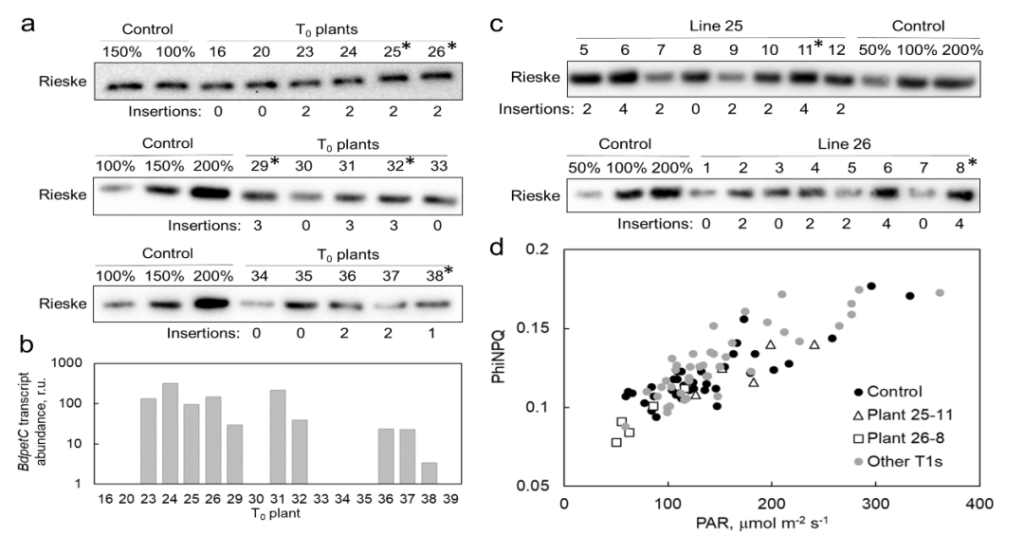
将这3个转基因株系的T2植株和对照植株,在自然光照射的温室中夏季种植。通过特异性抗体免疫印迹法,以叶面积为基础,分析叶片提取物中光合蛋白的丰度。结果显示,与对照植物相比,3个转基因株系的Rieske含量显著增加了约40%。而其他电子传递成分(如PSII的D1蛋白、ATP合成酶的AtpB亚基、PsbS和LHCII的Lhcb2亚基)的相对丰度和叶片Chl的相对含量基本没有改变。
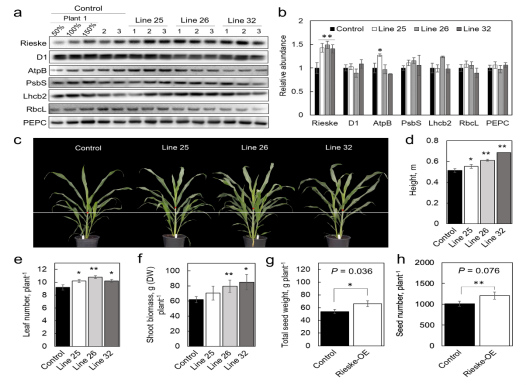
Rieske含量增加的高粱植株在发芽5周后比对照植株高,分蘖(0.036 < P > 0.09)和叶片更多。虽然基因型之间的叶片厚度和单位面积的叶片干质量没有差异,但收获时,品系26和32的地上总生物量高于对照植物。在另一项实验中,当植物在夏秋末在温室中种植时,25和26系的Rieske-OE植株的叶子比对照植物更大,并且比对照植物产生更多的种子。
为了了解过度表达Rieske的转基因高粱如何积累更多的生物量,研究人员分析了它们的光合特性。在恒定辐照度1500 μ mol m-2 s-1和不同CO2分压下,Rieske含量增加的植物与对照植物之间的CO2同化率或PSII (PhiPSII)的有效量子产率没有显著差异。在CO2环境下,在所有辐照度下,各基因型的CO2同化率和气孔导度相似。与对照植物相比,在不同辐照度下分析的PSI和PSII的光化学和非光化学产量在转基因植物中基本没有改变。这些结果表明,过表达Rieske植株的电子传递速率和CO2同化速率基本不受影响。而PSII的最大量子效率(FV/FM)在32系的植株中显著高于对照植株。
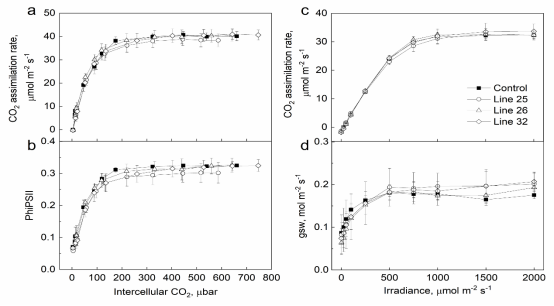
图3
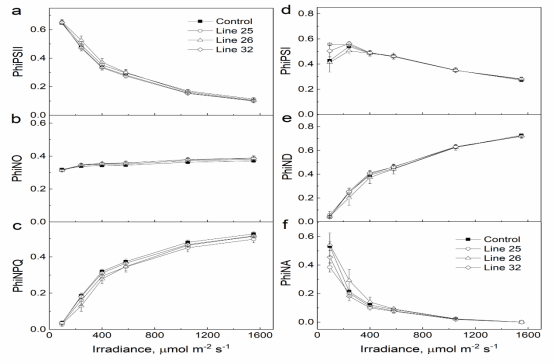
图4
在535 nm处,通过记录电致变色信号和吸光度变化来测试类囊体膜的通电性能,结果过表明过表达Rieske的植物与对照植物之间没有差异。为了获得qE以及ΔpH在光照过程中积累的动力学信息,研究人员记录了535 nm处的吸光度变化。过表达Rieske的转基因株系在从深色到1600 μ mol m-2 s-1的转变过程中建立qE的速度明显快于对照植物,表明ΔpH的建立速度更快,可能是由于Cytb6f活性的短暂增加引起的。
为了捕捉Cytb6f活性短暂增加对同化的影响,接下来分析了光合诱导,即光合作用在暗光转换时的激活过程,在夜间适应黑暗的植物在1000 μ mol m-2 s- 1光化光照射的前30分钟内。在诱导光合作用过程中,过表达Rieske的高粱植株CO2同化速率比对照植株加快,且在18 ~ 26 min内, 3个转基因株系的CO2同化速率均显著高于对照植株。PhiPSII的诱导动力学与同化动力学相似。与对照植物相比,转基因株系在光照开始后18 ~ 26min内更快达到稳态,25和26转基因系的PhiPSII显著增加。有趣的是,过表达Rieske的植株NPQ也表现出更动态的变化:在光照的前7min内NPQ的快速建立和NPQ的快速松弛。重要的是,尽管在光照开始后的16 ~ 23min内,过表达Rieske的3个株系的气孔导度都明显高于对照株系,但CO2利用率的增加并没有刺激CO2同化率的增加。在这段时间内,所有基因型的细胞间CO2分压与环境CO2分压的比值(Ci/Ca)都相似。
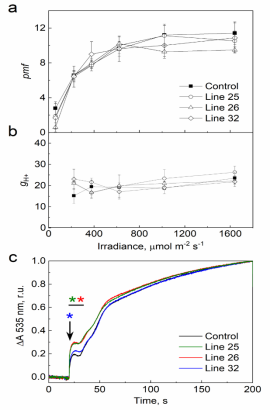
图5

综上所述:在高粱中,Rieske过表达对刺激电子传递的影响在光合作用诱导过程中更为明显,这表明其他光合成分共同限制了稳定的电子传递速率。然而,在过表达Rieske的暗适应高粱中,光合作用的更快激活导致了CO2同化率的短暂提高,并增加了生物量和粮食产量,这证实了光合作用的诱导限制了C4作物的生产力。因此,相对于其他电子传递成分,增加Rieske含量是提高C4作物生产力的一种有前途的方法,有助于确保未来的粮食和能源安全。
文章来源:植物生物技术Pbj