高蛋白的重要性?

图1 2022年11月15日,全球人口达到了80亿。图片来源:联合国人口基金。
1950年,世界人口总数约为26亿。到1987年,世界人口增长至50亿,1999年达到60亿,至2011年10月,全球人口约达70亿,而在上个月即2022年11月15日,我们一起见证了全球人口达到了80亿。预计,世界人口将从目前的80亿增至2050年的97亿,并可在2100年达到110亿的峰值。
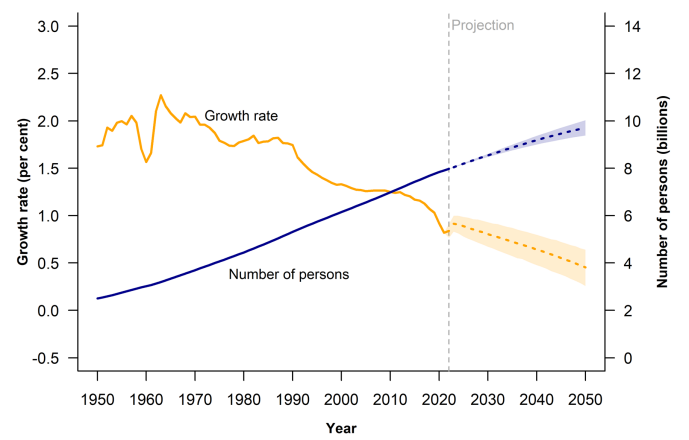
蛋白质已被确定为最受限制的宏量营养素,随着全球粮食需求的增加,将需要足够的蛋白质数量和质量(粮农组织,2013)。这么大的人口规模,意味着需要增加多少蛋白质?按目前的情况,我们能否满足?我们该做哪些努力?这些都是值得我们思考的问题。
玉米籽粒被认为可以将干物质转化为肉、奶和蛋的效率最高,因此被赋予“饲料大王”的称号,被称为最佳的能量饲料。
虽然玉米被世界公认为是饲养家禽的首选谷物,但它仍有不足的地方,主要表现在其蛋白质含量低、蛋白质质量差,因此需要在玉米饲料中使用高蛋白补充剂或添加额外的氨基酸如赖氨酸、色氨酸,比如可以和豆类适当混合来克服。
玉米籽粒中各种营养素的浓度取决于基因型、农艺管理、基因型与环境之间的相互作用以及收获后处理,玉米的遗传变异很大,可以通过开发某些营养素更高的改良品种来改善目前的状况 (Dei, 2017)。
曾经没人做过吗?做了。例如,通过育种开发的玉米品种含有高达21%的粗蛋白,然而粗蛋白含量超过12%的玉米品种的产量确实较低 (Boyat, 1980),因此不适合商业化生产。可见,失败了。
所以说,不定位出高蛋白基因,是很难实现这个伟大的愿景的。为什么几十年来没人定位出来,很难定位吗?这与它是数量性状基因有关,就同定位植物的高度、产量这类基因一样,如大海捞针,可以想象难度非常大!
表1 正常玉米籽粒和籽粒部分的化学成分(%DM)(Dei, 2017)。
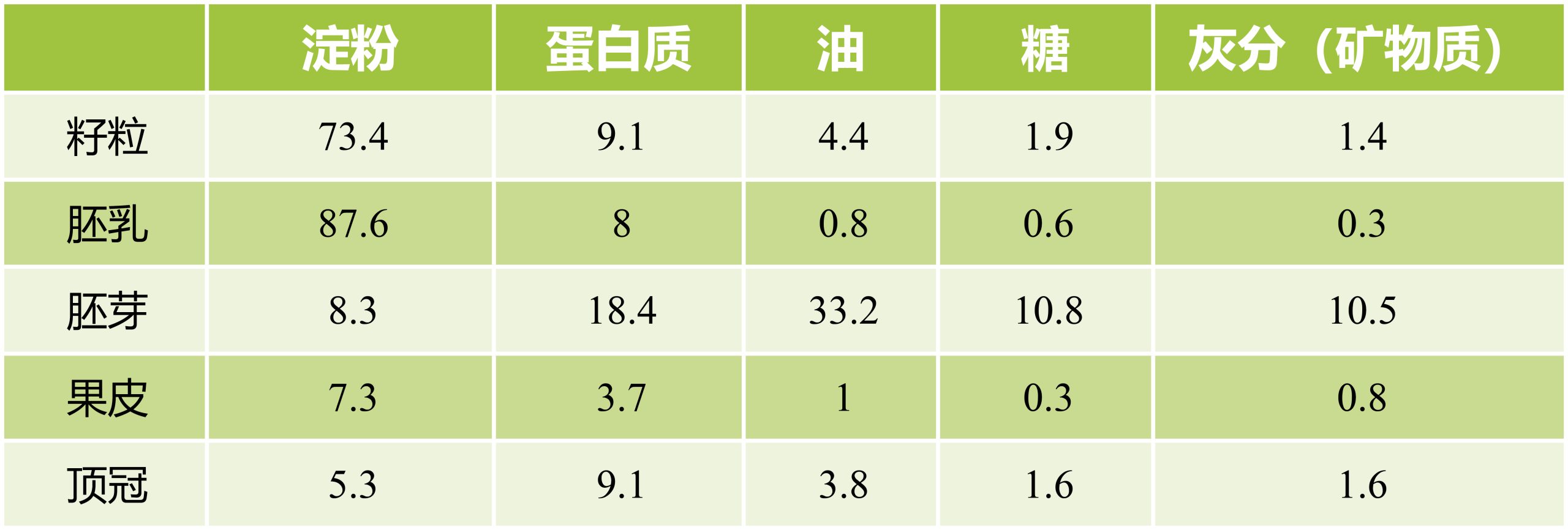

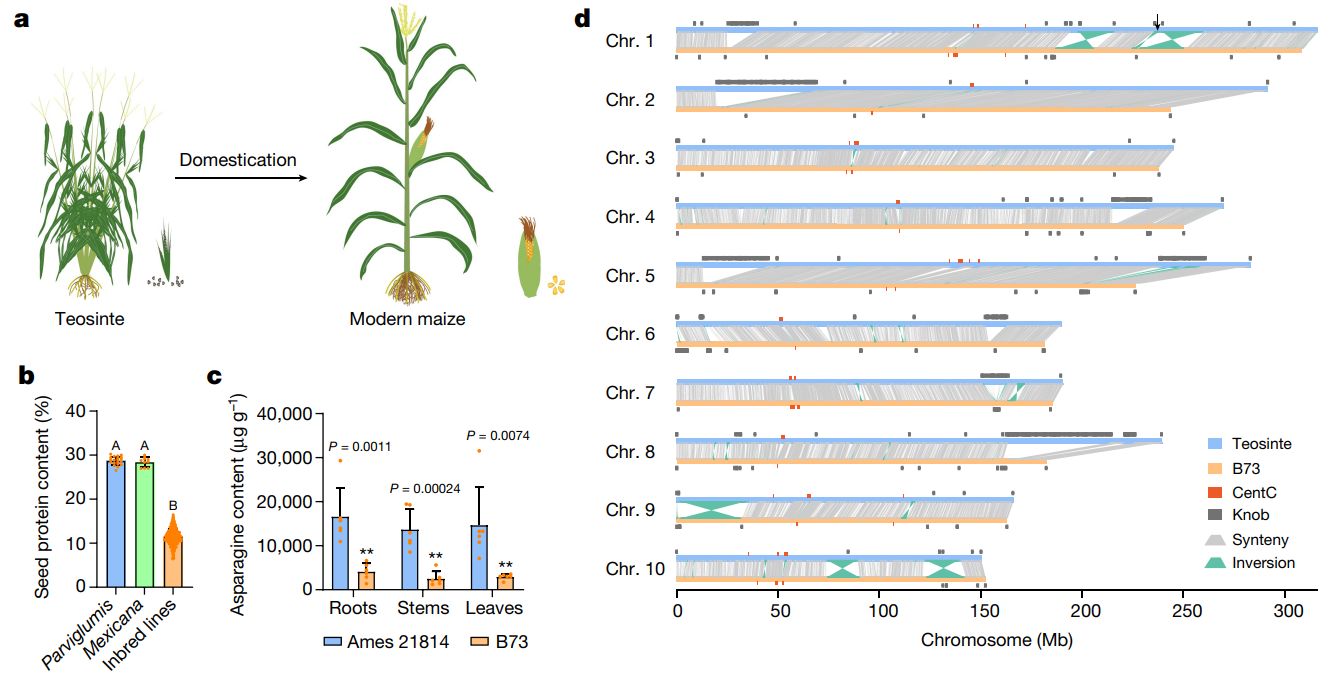
图3 大刍草和现代玉米之间蛋白质含量、基因序列的比较。(a)大刍草和现代玉米的示意图;(b)比较各材料籽粒中的蛋白质含量;(c)Ames 21814和B73在根、茎和叶中游离天冬酰胺的含量比较;(d)B73参考基因组(B73_v5)(橙色)和Ames 21814(蓝色)之间的共线性分析。
实验:收集玉米种质资源、检测各材料的蛋白质含量。
结果:收集了20份大刍草(Zea mays subsp. parviglumis)、10份墨西哥大刍草(Zea mays subsp. mexicana)和518份玉米自交系;在大刍草各品系材料中蛋白质含量均在30%左右,而在405份最终被检测的玉米自交系中,蛋白质含量在6.5%-16.5%之间,均值为11.5%(图3b)。
结论:大刍草和现代玉米自交系的蛋白质含量存在显著性差异,可能存在某些基因位点没有被选择导致了这两者的差异,这些位点在自交系中是遗传可变的。
2,比较Ames 21814和B73的蛋白质含量及氨基酸组成
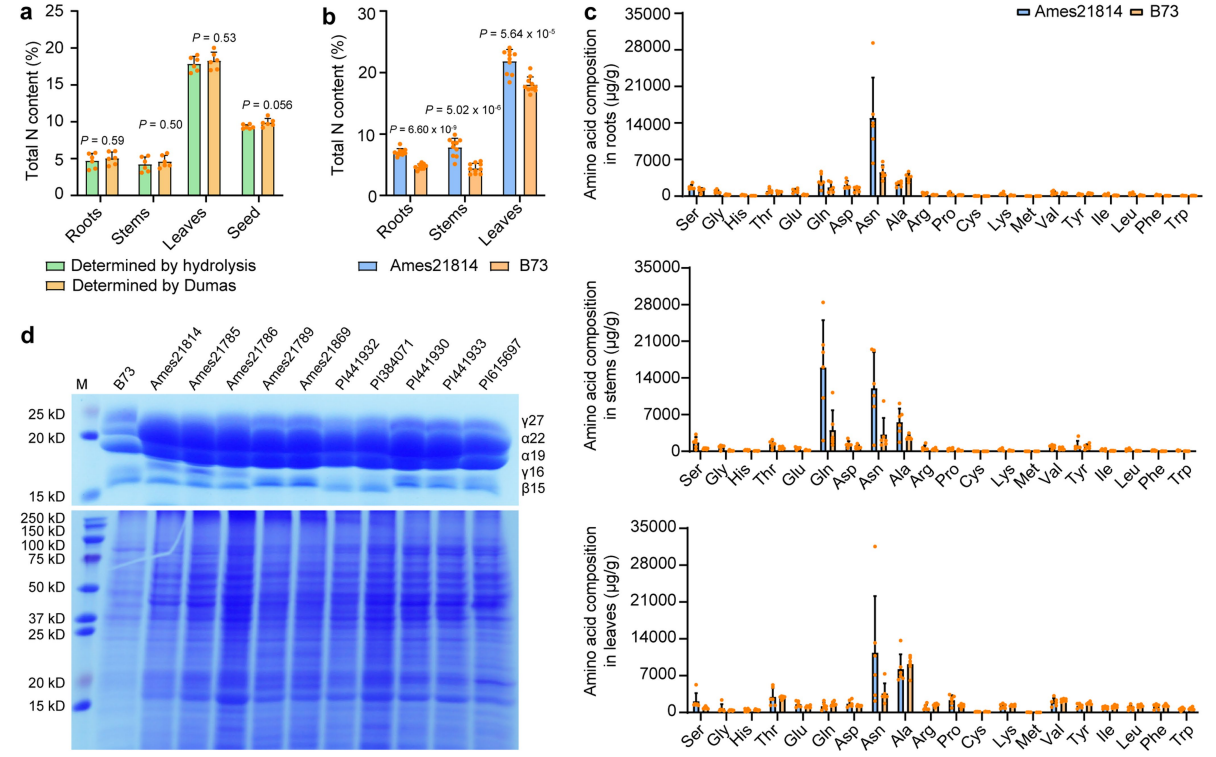
图4 Ames 21814和B73在根、茎和叶中的总氮量、游离氨基酸含量的比较,以及对它们的玉米醇溶蛋白和非醇溶蛋白进行SDS-PAGE实验。(a)通过酸水解和Dumas两种方法检测B73在根、茎和叶中的总氮量;(b)Ames 21814和B73在根、茎和叶中的总氮量比较;(c)Ames 21814和B73在根、茎和叶中的游离氨基酸含量的比较;(d)对B73和10株大刍草的玉米醇溶蛋白和非醇溶蛋白进行SDS-PAGE实验。
实验:检测Ames 21814和B73不同部位的蛋白质含量、游离氨基酸含量;比较Ames 21814和B73籽粒中玉米醇溶蛋白与非醇溶蛋白含量。
结果:检测根、茎、叶中的蛋白质含量,Ames 21814均比B73有显著性增加;检测根、茎、叶中的游离氨基酸含量,最显著的差异在于Ames 21814中的天冬酰胺含量比B73高(图4c);与B73相比,大刍草籽粒中的玉米醇溶蛋白尤其是α19和α22有显著积累(图4d)。
结论:天冬酰胺水平的增加与高蛋白质含量有关。
备注:玉米籽粒中的蛋白按照其可溶性分为玉米醇溶蛋白(prolamines/zeins)、白蛋白(albumins)、球蛋白(globulins)和谷蛋白(glutelins)。玉米醇溶蛋白是主要的胚乳贮藏蛋白,占比60%以上,其包括四个家族:α(19-和22-kDa,分别命名为α19、α22)、β(15-kDa)、γ(50-,27-和16-kDa)和δ(18-和10-kDa)。
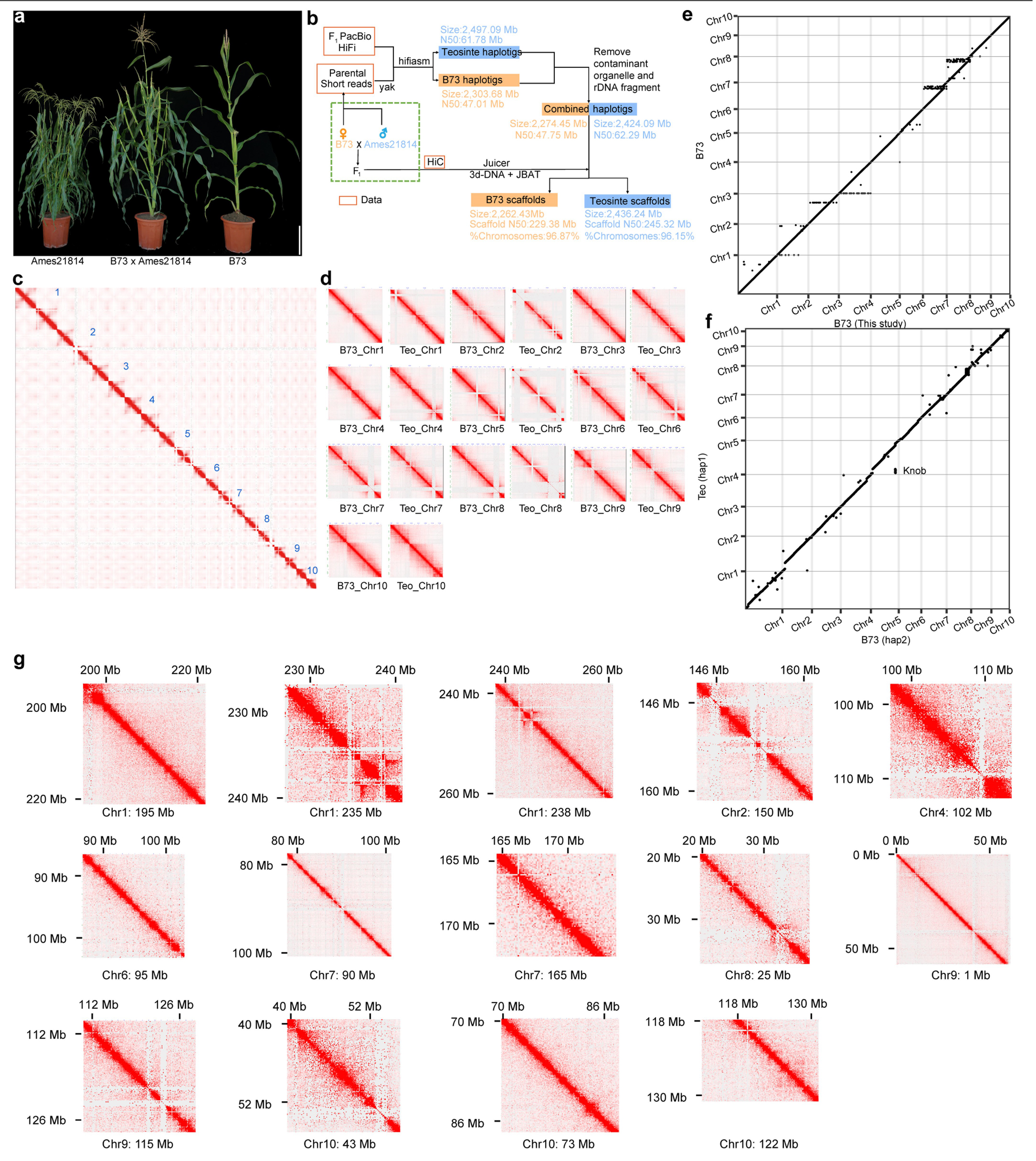
图5 使用Trio binning对大刍草Ames 21814基因组进行单倍型组装。
实验:HiFi和HiC测序,Trio binning组装。
结果:将B73×Ames 21814的F1单株进行测序,利用测序得到的47×深度的104Gb HiFi数据,组装得到了2424Mb的Ames 21814基因组, contig N50为62.29Mb。将375.56Gb Hi-C数据锚定到10条染色体上(图5d),最终得到2436Mb大小的高质量Ames 21814基因组,其中包括452 scaffolds、245.33Mb N50 scaffold。
尽管Ames 21814和B73_v5有很好的共线性,但仍包含了280881个结构性变异,其中大于100kb的倒位有71个。
表2 大刍草Ames 21814基因组的组装结果。

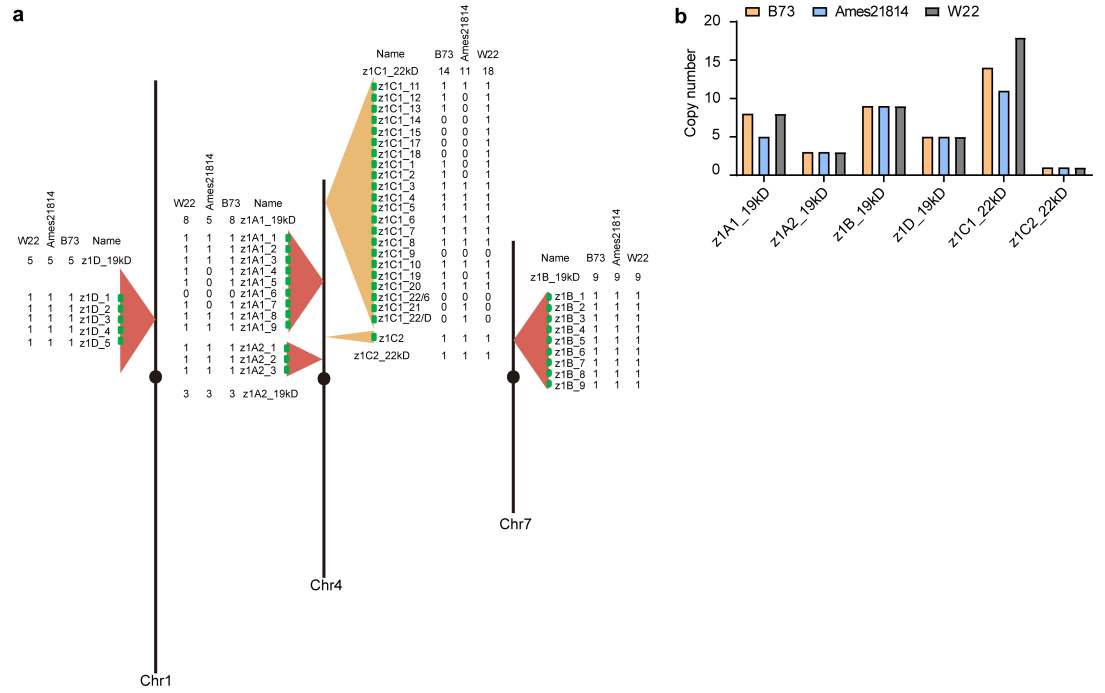
图6 在Ames 21814和玉米自交系中,对α型玉米醇溶基因的拷贝数进行比较。(a)Ames 21814、B73和W22中α型玉米醇溶基因的标注示意图;(b)对拷贝数进行统计分析。
在Ames 21814中,分别有22个α19和12个α22拷贝数;在B73中,分别有25个α19和15个α22拷贝数(图6)。
结论:是否是因为结构变异导致大刍草和玉米自交系的表型差异仍需要通过实验来进行验证。大刍草的高蛋白性状并不是因为它含有更多的α型玉米醇溶基因拷贝数,导致高蛋白性状的QTL另有其他位点。
4,克隆高蛋白基因
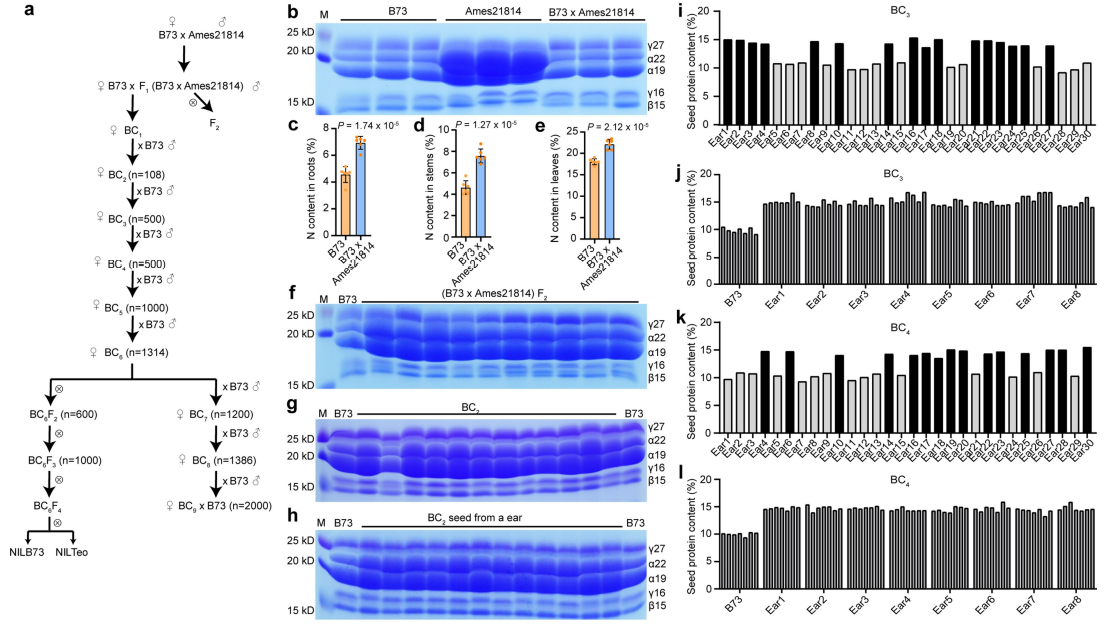
图7 遗传群体的创建以及蛋白质含量的测定。(a)遗传群体的创建过程,群体大小已标注;(b)SDS-PAGE检测B73、Ames 21814、B73×Ames 21814籽粒中的玉米醇溶蛋白;(c-e)B73、B73×Ames 21814的F1植株的根、茎和叶的总氮量;(f-h)SDS-PAGE检测B73×Ames 21814的F2籽粒、BC2果穗、BC2籽粒中的醇溶蛋白;(i)检测来自30个不同的BC3果穗的籽粒中的蛋白质含量;(j)BC3群体中8个高蛋白质含量的果穗的蛋白质含量;(k)检测来自30个不同的BC4果穗的籽粒中的蛋白质含量;(l)BC4群体中8个高蛋白质含量的果穗的蛋白质含量。
备注:“果穗”其实就是指“玉米棒子”喔!
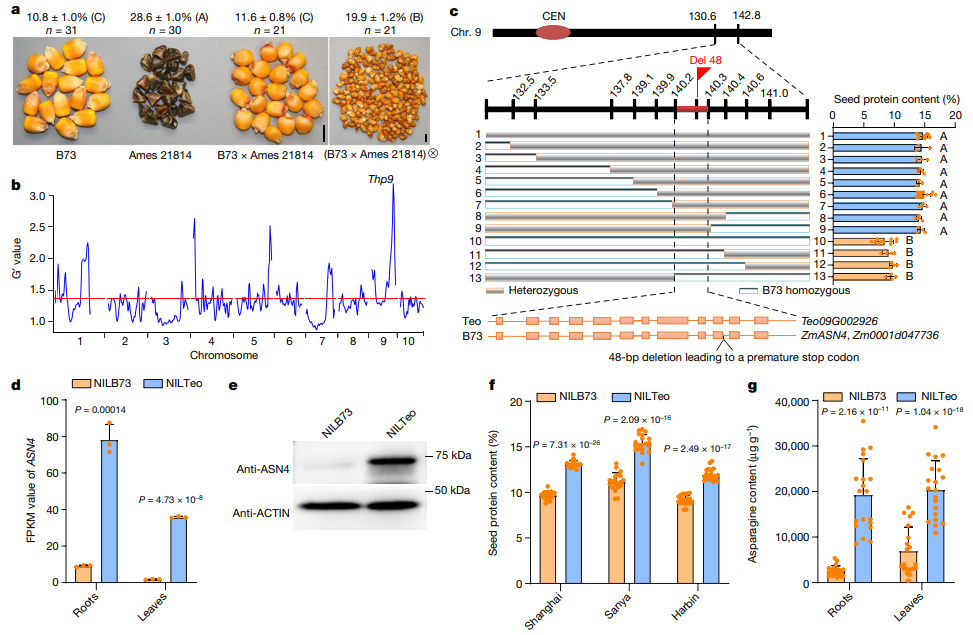
图8 对THP9的图位克隆。(a)B73、Ames 21814、B73×Ames 21814及其自交后代的籽粒表型,上面的数字指籽粒中的蛋白质含量;(b)对BC4群体的BSA-seq分析;(c)THP9的图位克隆,THP9位于140.2和140.3两个分子标记之间150kb的位置;(d)THP9在NILTeo和NILB73中的FPKM值;(e)免疫印迹检测NILTeo和NILB73根中的ASN4蛋白;(f)检测在上海、三亚和哈尔滨收获的NILTeo和NILB73籽粒的蛋白质含量;(g)检测NILTeo和NILB73根中的游离天冬酰胺含量。
实验:建立遗传群体,BSA-seq,重测序,RNA-seq,双荧光素酶报告基因实验。
结果:
以Ames 21814为高蛋白供体亲本,以B73为反复回交亲本,建立连续回交群体(图7a)。F1籽粒的蛋白质含量(11.6±0.8%)类似于B73(10.8±1.0%),而Ames 21814籽粒的蛋白质含量为28.6±1.0%(图8a),这个结果也与玉米醇溶蛋白在各品种中的积累模式相符,因为如前所述,α型玉米醇溶蛋白是总蛋白质含量的标志物(图7b)。F1植株根、叶的总氮量高于B73(图7c-e),F2籽粒的蛋白质含量(19.9±1.2%)几乎是F1和B73籽粒的两倍(图8a)。
种植BC3籽粒,从每株植物中保存一片叶子用于DNA提取,汇集低蛋白和高蛋白的DNA样本(每种n=75)用于BSA-seq。结果显示,在1、3、4、5、7和9号染色体上有几个QTLs位点,在9号染色体130Mb-160Mb之间有一个显著的峰值,包含了315个大刍草基因,该位点被命名为THP9。
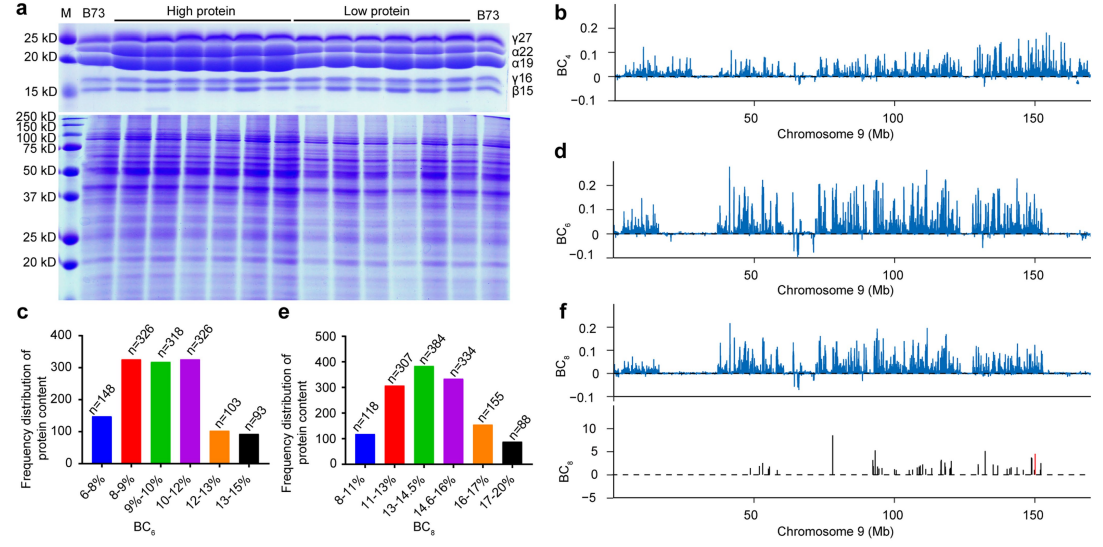
图9 对BC4、BC6和BC8三个群体进行BSA-seq以定位THP9,并对5个高蛋白质含量和5个低蛋白质含量的BC6F4株系进行深度重测序。(a)通过对BC4群体中的果穗进行玉米醇溶蛋白(上图)和非醇溶蛋白(下图)检测来进行高、低蛋白含量的表型分析;(b)基于BSA-seq,对BC4群体进行基因渗入分析;(c)对BC6群体的籽粒蛋白质含量进行统计分析;(d)基于BSA-seq,对BC6群体进行基因渗入分析;(e)对BC8群体的果穗的蛋白质含量进行频率分布分析;(f)基于BSA-seq,对BC8群体进行基因渗入分析;(g)对10个株系进行重测序分析,将候选区域缩小至135.5-143Mb之间。
创建BC6(n=1314)和BC8(n=1344)群体,同上,对其进行BSA-seq,同样证实了THP9位点的存在,分别包含271个和190个大刍草基因(图9c-f)。对BC6F4群体中选取5个高蛋白质含量和5个低蛋白质含量的株系进行深度重测序(大于20×),最小的共同区域135.5-143Mb称为候选区域(图9g),并在此区间上建立了高蛋白质含量和低蛋白质含量的近等基因系NILTeo和NILB73。
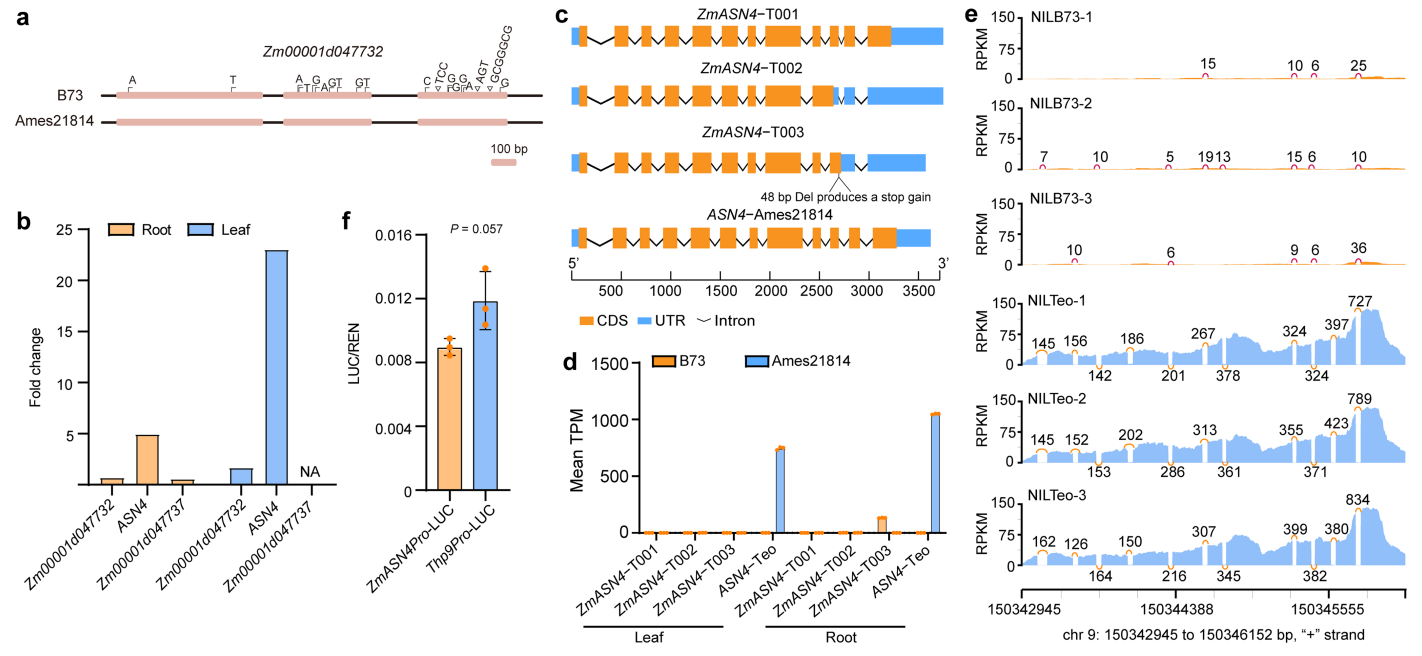
继续创建BC9群体(n=2000),将THP9缩小至一个150kb的区域,该区域包含三个候选基因,Zm00001d047732、Zm00001d047736和Zm00001d047737。经分析及检测后发现,Zm00001d047736对应Ames 21814中的Teo09G002926,写为THP9-Teosinte或THP9-T,其编码天冬酰胺合成酶4蛋白(ZmASN4),而Zm00001d047736在ASN4的第10个内含子缺失48bp,写为THP9-B73或THP9-B(图8c)。
THP9-B中内含子的缺失使其产生了三种不同的转录本,ZmASN4-T001与THP9-T(也可记为ASN4-T)类似,而ZmASN4-T002、ZmASN4-T003均包含了一个过早的终止密码子(图10c),但这三种转录本在B73中几乎检测不到(图10d)。
利用双荧光素酶报告基因实验比较THP9-T和THP9-B的启动子活性,结果显示它们无显著性差异(图10f),说明这两个基因的差异表达并不是由启动子变异引起的。与转录水平一致,ASN4蛋白在NILTeo中大量积累,而在NILB73中无法检测到(图8e)。这些结果表明,B73中ASN4基因48bp的缺失显著影响了其转录本的RNA剪接和稳定性,使转录本和ASN4蛋白均难以被检测到。THP9-B可以被认为是一个空等位基因。
利用分子标记对BC7群体进行基因分型,检测到基因型为THP9-T和THP9-H(T/B)的植株的根中,游离天冬酰胺的含量显著高于基因型为THP9-B的植株的根。基因型为THP9-T和THP9-H(T/B)的植株的籽粒中,蛋白质含量也显著高于基因型为THP9-B的植株的籽粒(图11b、c)。
在哈尔滨、上海、三亚分别种植的NILTeo籽粒的蛋白质含量分别为12±0.7%、13.1±0.4%和15.4±1.0%,而对应地区的NILB73籽粒的蛋白质含量分别为9.2±0.5%、9.7±0.4%和11.2±1.0%。可见,THP9-T等位基因使这三个地方的玉米蛋白质含量分别增加了30.4%、35.2%和37.8%。对于其他表型,与NILB73相比,NILTeo株高增加了7.6%,植物鲜重增加了15.1%(图11e、f)。
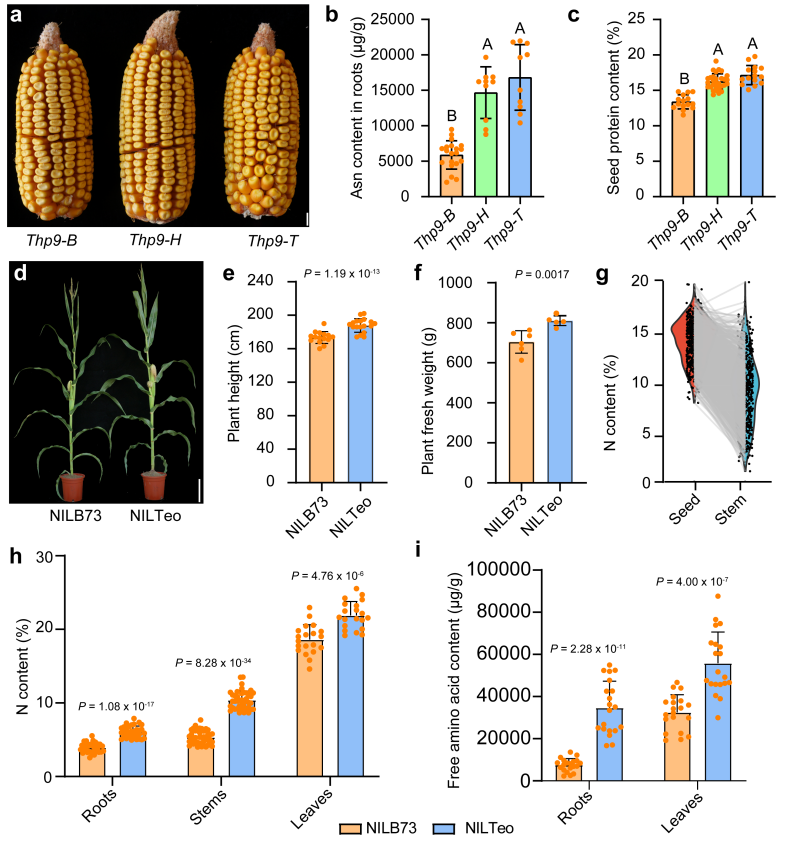
图11 THP9-T与BC7F3群体中高天冬酰胺和高蛋白含量的关系以及NILTeo和NILB73的表型比较。(a)三种基因型——纯合THP9-B、杂合THP9-H(T/B)和纯合THP9-T玉米果穗的代表性表型照片;(b)BC7F3群体中THP9-B、THP9-H和THP9-T根中游离天冬酰胺的含量;(c)BC7F3群体中THP9-B、THP9-H和THP9-T自交后收获的籽粒的蛋白质含量;(d)近等基因系NILTeo和NILB73的表型照片;(e)NILTeo和NILB73的株高比较;(f)NILTeo和NILB73的鲜重比较;(g)BC8群体籽粒与茎总氮量的关联分析;(h)检测NILTeo和NILB73的根、茎和叶的总氮量;(i)检测NILTeo和NILB73根、叶中的游离氨基酸含量。
使用快速氮分析仪对1334株BC8群体的茎和果穗进行检测,结果显示它们高度相关(图11g)。在NILTeo中,其根、茎和叶组织的总氮量与游离氨基酸含量均显著高于NILB73(图11h、i),其根中的天冬酰胺含量显著高于NILB73(图8g)。
结论:THP9-B第10个内含子缺失48bp导致其转录本和ASN4蛋白几乎检测不到;THP9-T等位基因与更高的蛋白质含量相关;THP9-T通过增加天冬酰胺的积累促进了根、茎、叶和籽粒中蛋白质含量的增加。
5,对THP9的验证和自然变异的研究
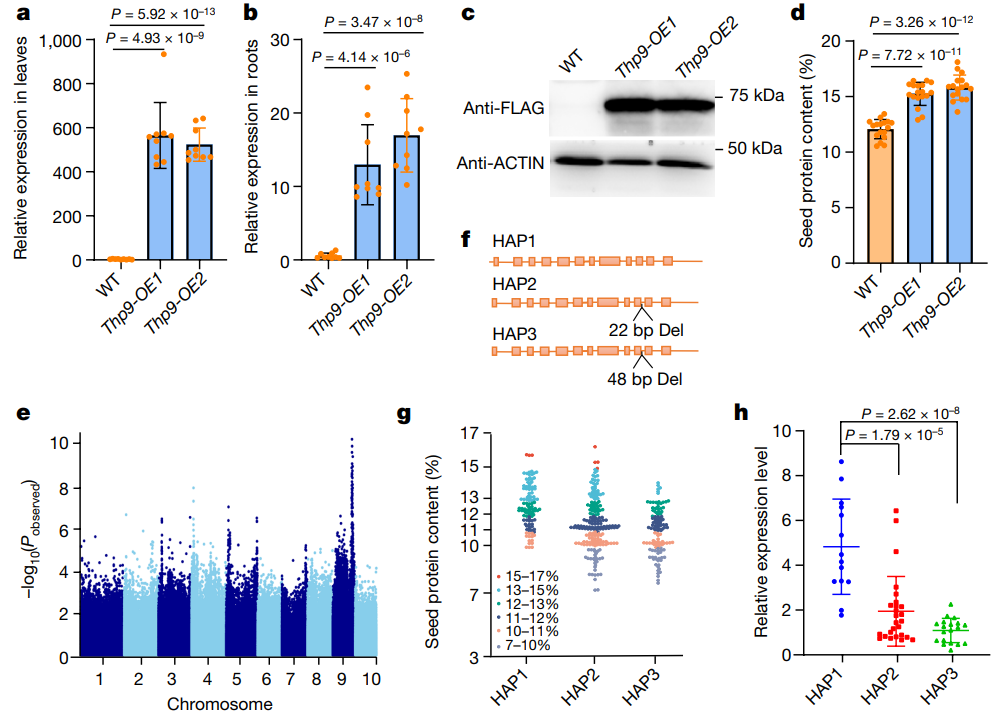
图12 对THP9的验证和自然变异的研究。(a、b)qRT-PCR检测THP9在THP9-OE1和THP9-OE2叶、根中的表达情况;(c)检测在THP9-OE1和THP9-OE2叶中ASN4的蛋白表达情况;(d)检测THP9-OE1和THP9-OE2籽粒中蛋白质含量;(e)与玉米自交系籽粒蛋白含量相关联的GWAS分析;(f)ASN4基因有三种基因型:HAP1、HAP2和HAP3;(g)检测HAP1、HAP2和HAP3籽粒的蛋白含量;(h)检测HAP1、HAP2和HAP3根中ASN4基因的表达情况。
实验:转基因验证实验,全基因组关联研究(GWAS),蛋白质含量测定,荧光定量PCR。
结果:
与非转基因B73植株相比,过表达THP9-T基因的植株的叶片、根中ASN4在转录水平、蛋白水平有显著升高(图12a-c)。两个过表达株系THP9-OE1和THP9-OE2的籽粒中,蛋白质含量分别为15.2±1%和15.8±1.1%,分别比B73对照(12.1±0.9%)增加了25.7%和30.9%(图12d)。
2019年测量了405个玉米自交系的籽粒蛋白质含量,2020年测量了437个玉米自交系的籽粒蛋白质含量,利用这些蛋白质含量数据进行GWAS分析,确定了THP9位点附近的一个物理坐标区域(图12e)。对420个自交系的PCR及测序发现,ASN4基因的第10个内含子有三种基因型(HAP1-HAP3):占比25.7%的HAP1自交系具有Ames 21814基因型(THP9-T),ASN4编码序列完整;占比46.4%的HAP2自交系在第10个内含子中缺失22bp(记为THP9-T’);占比27.9%的HAP3自交系具有B73基因型(THP9-B),内含子缺失48bp(图12f)。
将蛋白质含量高于13%的株系定义为高蛋白自交系,蛋白质含量低于10%的株系定义为低蛋白自交系。HAP1自交系的平均蛋白质含量最高(12.7±1.4%),HAP2自交系的平均蛋白质含量中等(11.5±1.7%),HAP3自交系的平均蛋白质含量最低(11.0±1.4%)(图12g)。qRT-PCR结果显示,HAP1中ASN4的转录水平普遍高于HAP2和HAP3型(图12h),与蛋白质含量正相关。
结论:转基因株系的检测结果与THP9-B导致B73低蛋白质含量的假设一致。ASN4基因的第10个内含子的三种基因型表明内含子缺失的长度与最终转录水平有关。以上结果也支持了THP9是影响自交系籽粒蛋白质含量变化的主要QTL的假设。
6,THP9-T提高了氮素利用效率(NUE)
实验:外源氮源对植株生长的影响,蛋白质含量测定。
结果:
在上海修建了含不同氮浓度土壤的容器(图13a-d)。正常施氮条件(每株施用40g)和低施氮条件(不施用氮)下,NILTeo植株比NILB73植株生长的更好(图13f-h)。
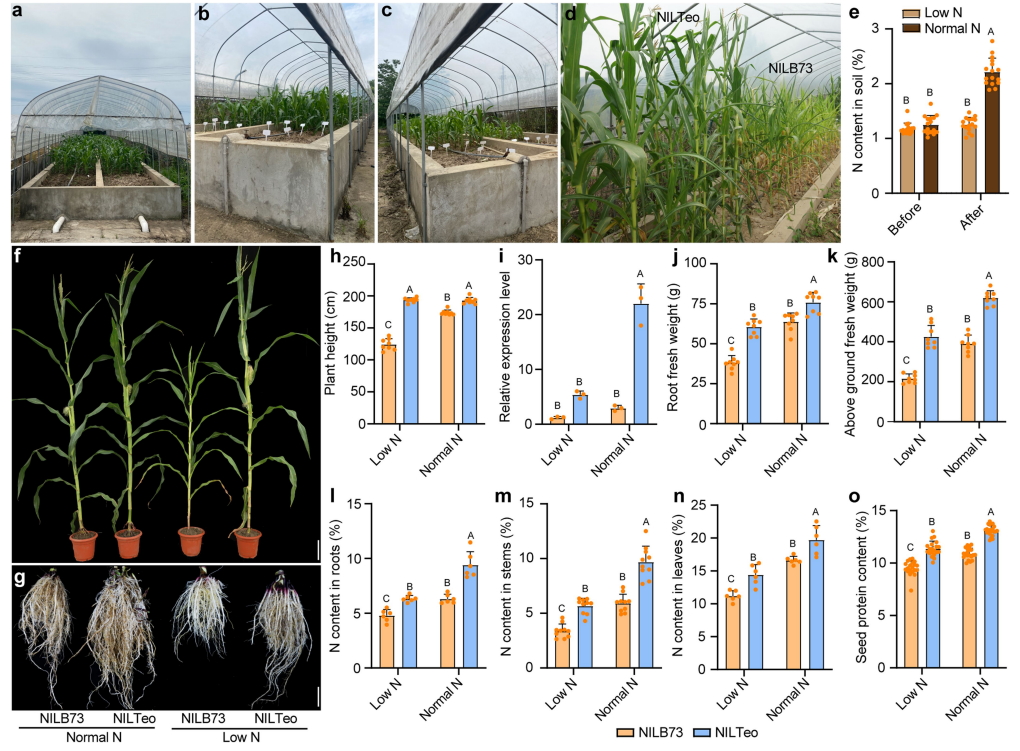
图13 NILB73和NILTeo在正常施氮条件和低施氮条件下的比较。(a-c)修建的含不同施氮浓度土壤的容器;(d)不施氮情况下,NILB73和NILTeo的生长情况;(e)不同施氮条件下,种植前后土壤中的氮含量;(f、g)NILB73和NILTeo在不同施氮条件下的植株表型及根部表型;(h)不同施氮条件下,NILB73和NILTeo的株高比较;(i)不同施氮条件下,NILB73和NILTeo的根中THP9的表达情况;(j)不同施氮条件下,NILB73和NILTeo根的鲜重比较;(k)不同施氮条件下,NILB73和NILTeo地上生物量的比较;(l-n)不同施氮条件下,NILB73和NILTeo根、茎和叶的总氮量比较;(o)不同施氮条件下,NILB73和NILTeo籽粒蛋白质含量的比较。
qRT-PCR结果显示,施氮后对THP9-T的诱导表达作用较强,而THP9-B不表达(图13i),说明THP9-T对土壤氮水平敏感。在低施氮条件下,与NILTeo相比,NILB73植株在根鲜重、地上生物量上显著降低(图13j、k),THP9-OE2也表现出类似的现象(图14a-j)。
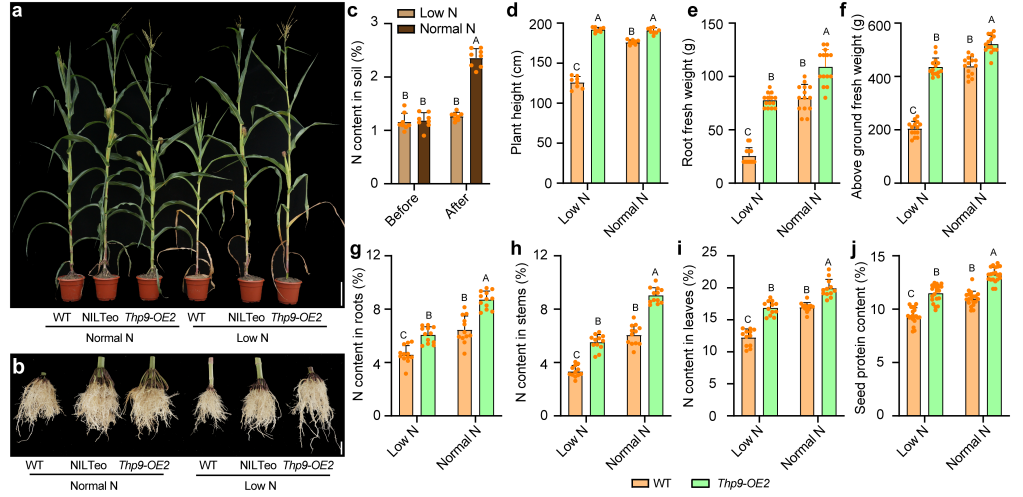
图14 THP9-OE2和B73在正常施氮条件和低施氮条件下的比较。(a)不同施氮条件下,B73、NILTeo和THP9-OE2的表型比较;(b)不同施氮条件下,B73、NILTeo和THP9-OE2的根部表型比较;(c)不同施氮条件下,种植前后土壤中的氮含量;(d)不同施氮条件下,B73和THP9-OE2的株高比较;(e)不同施氮条件下,B73和THP9-OE2根的鲜重比较;(f)不同施氮条件下,B73和THP9-OE2地上生物量的比较;(g-i)不同施氮条件下,B73和THP9-OE2根、茎和叶的总氮量比较;(j)不同施氮条件下,B73和THP9-OE2籽粒蛋白质含量的比较。
在三亚进行了更大的田间实验(图15a),设置了四种不同浓度的施氮量:100%(每株施用32g)、50%(每株施用16g)、25%(每株施用8g)和0%。结果显示,在不同施氮浓度条件下,NILTeo在株高(图15b)和地上生物量(图15c)方面均比NILB73更有生长优势。在所有实验中,NILTeo的根、茎、叶的总氮含量均显著高于NILB73(图15d-f)。将施氮量从100%降至0%后,NILTeo籽粒的蛋白质含量从14.2%降至13.5%、12.0%和10.7%,而NILB73籽粒的蛋白质含量从11.4%降至11.2%、10.7%和8.9%(图15g)。在每次处理中,NILTeo籽粒的蛋白质含量始终高于NILB73。在25%施氮条件下收获的NILTeo籽粒蛋白质含量为12.0%,高于正常施氮条件下收获的NILB73籽粒(11.4%)。
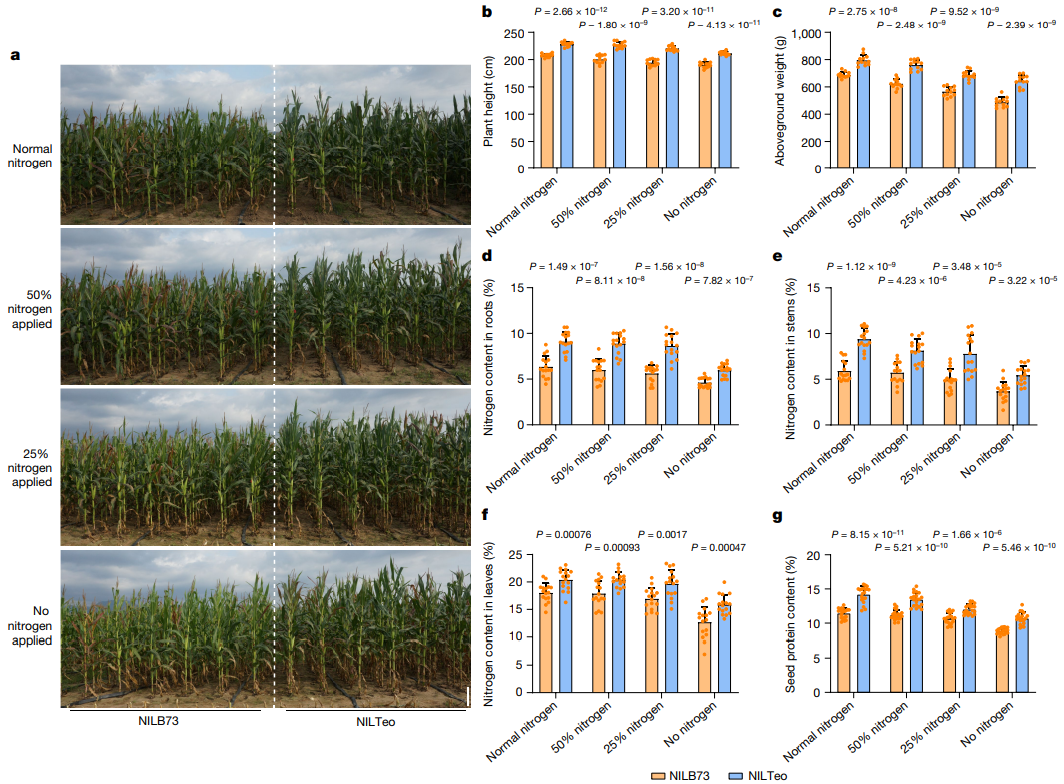
图15 NILTeo和NILB73的NUE比较。(a)四种施氮条件下,NILTeo和NILB73的生长情况;(b)四种施氮条件下,NILTeo和NILB73的株高比较;(c)四种施氮条件下,NILTeo和NILB73的地上生物量的比较;(d-f)四种施氮条件下,NILTeo和NILB73的根、茎和叶的总氮量比较;(g)四种施氮条件下,NILTeo和NILB73的籽粒蛋白质含量的比较。
结论:在正常和低施氮条件下,NILTeo比NILB73表现出更高的NUE,THP9-OE2比B73表现出更高的NUE。
7,利用THP9-T进行玉米种质创新
实验:杂交实验,蛋白质含量测定。
结果:
收获NILTeo×Mo17和NILB73×Mo17的F2果穗,检测结果显示,NILTeo×Mo17籽粒的蛋白质含量(9.2±0.6%)比NILB73×Mo17籽粒的蛋白质含量(8.6±0.4%)高7.8%,而两者的杂交种的百粒重几乎相同(图16a-c)。

将THP9-T导入郑单958(记为郑单958-T),与郑单958(也记为郑单958-B)相比(图16d、e),郑单958-T籽粒的蛋白质含量为11.1±1.1%,比郑单958-B增加了12.7%(图16h),其根、茎和叶的总氮含量也显著增加(图16i-k),但百粒重无显著差异(图16l)。
结论:THP9-T具有改善玉米种质资源的潜力。
将THP9-T转入玉米自交系中,使种子蛋白质含量增加约35%,根中氮含量增加约54%,茎中氮含量增加约94%,叶片中氮含量增加约 18%,并且生物量也大大增加。并且在低氮条件下,表现出更高的氮素利用效率。将其导入我们推广面积最大的玉米杂交种郑单958中,可显著增加籽粒的蛋白质含量。这将有利于普通栽培玉米籽粒中蛋白质含量的遗传改良,并对减少化肥使用和保护生态环境具有很好的指导意义。
United Nations, “World Population Prospects,” 2022, http://www.un.org/.
Boyat M, Deriux M, Kaan F, Rautou S. Maize breeding for improvement of kernel protein content using Illinois high protein population. In: Pollmer WG, editor. Improvement of Quality Traits of Maize for Grain and Silage. Pflanzenzuchtag. 1980; 80: 173–183.
Dei, H. K. (2017). Assessment of Maize (Zea mays) as Feed Resource for Poultry. In (Ed.), Poultry Science. IntechOpen. https://doi.org/10.5772/65363
Huang Y, Wang H, Zhu Y, et al. THP9 enhances seed protein content and nitrogen-use efficiency in maize [published online ahead of print, 2022 Nov 16]. Nature. 2022;10.1038/s41586-022-05441-2. doi:10.1038/s41586-022-05441-2
Lonnie M, Hooker E, Brunstrom JM, et al. Protein for Life: Review of Optimal Protein Intake, Sustainable Dietary Sources and the Effect on Appetite in Ageing Adults. Nutrients. 2018;10(3):360. Published 2018 Mar 16. doi:10.3390/nu10030360
Prasanna BM, Palacios-Rojas N, Hossain F, et al. Molecular Breeding for Nutritionally Enriched Maize: Status and Prospects. Front Genet. 2020;10:1392. Published 2020 Feb 21. doi:10.3389/fgene.2019.01392
