
种子休眠是是指种子在适合它生长的条件(温度、水分和氧气等)下仍不能发芽的现象,是多数高等植物所共有的特点。作物驯化过程中由于更多考虑高产、优质、抗病虫及耐受逆境性状,同时保证在生产中种子具有一致的萌发特性,常常忽视了对种子适度休眠的保留,从而导致很多作物如水稻、小麦会发生穗发芽现象(种子收获前成熟期如遇连阴雨不能及时收获,常出现部分籽粒在穗上发芽的现象),严重影响了作物的产量和品质,造成巨大的经济损失(图1)。仅在面包小麦中,每年由于穗发芽造成的损失就要高达10亿美元。在中国南方水稻栽培区,受收获季节梅雨的影响,穗发芽会造成常规稻6%栽培面积的损失,而杂交稻的损失则高达栽培面积的20%。近年来,随着全球气候变暖,农业生产上水稻小麦等在后期成熟期常常遭遇连阴天气,使得穗发芽灾害频繁发生。如我国长江中下游以及黄淮地区等冬小麦主产区在2013、2015及2016年均遭受严重的穗发芽灾害。在2016和2020年,受台风影响,江浙地区水稻、玉米等也爆发大面积穗发芽,造成严重减产。更为严重的是,在作物制种后期如发生连绵阴雨天气,引起穗发芽将对制种产业造成难以估量的损失,甚至影响下季播种。因此,找到水稻、小麦等控制种子休眠的关键基因,阐明种子休眠调控的分子生理机制,挖掘其优良等位变异,对解决水稻等作物穗发芽灾害至关重要。

图 1. 2020年收获季连绵阴雨导致南方不同省份水稻穗发芽
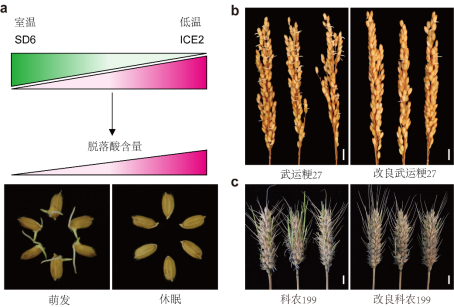
a, SD6/ICE2分子模块作用模型示意图;b, 改良前后水稻品种穗发芽情况;c, 改良前后小麦品种穗发芽情况。
Fang, J., Chai, C., Qian, Q., Li, C., Tang, J., Sun, L., Huang, Z., Guo, X., Sun, C., Liu, M., Zhang, Y., Lu, Q., Wang, Y., Lu, C., Han, B., Chen, F., Cheng, Z., and Chu, C. (2008) Mutations of genes in synthesis of the carotenoid precursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice, The Plant Journal. 54, 177–189.
Fang, J. and Chu, C. (2008). Abscisic acid and the pre-harvest sprouting in cereals. Plant Signal and Behavior. 3: 1046-1048.
Du, L., Xu, F., Fang, J., Gao, S., Tang, J., Fang, S., Wang, H., Tong, H., Cao, S., Zhang, F., Chu, J., Wang, G., and Chu, C. (2018). Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice. The Plant Journal. 95, 545–556.
Wang, M., Li, W., Fang, C., Xu, F., Liu, Y., Wang, Z., Yang, R., Zhang, M., Liu, S., Lu, S., Lin, T., Tang, J., Wang, Y., Wang, H., Lin, H., Zhu, B., Chen, M., Kong, F., Liu, B., Zeng, D., Jackson, S.A.*, Chu, C.*, and Tian, Z.* (2018). Parallel selection on a dormancy gene during domestication of crops from multiple families. Nature Genetics. 50, 1435–1441.
Xu, F., Tang, J., Gao, S., Cheng, X., Du, L., and Chu, C. (2019). Control of rice pre-harvest sprouting by glutaredoxin-mediated abscisic acid signaling. The Plant Journal. 100, 1036–1051.
Xu, F.#, Tang, J.#, Wang, S.#, Cheng, X.#, Wang, H., Ou, S., Gao, S., Li, B., Qian, Y., Gao, C.*, and Chu, C.* (2022). Antagonistic control of rice seed dormancy by two bHLH transcription factors. Nature Genetics. doi: 10.1038/s41588-022-01240-7.