通过植物育种提高作物产量是费时费力的,新的等位基因组合的产生受到染色体连锁的限制。减数分裂重组是通过重组亲本等位基因创造新的遗传变异。同源染色体之间的遗传信息交换发生在交叉位点(CO)上,但交叉位点的频率通常较低且分布不均匀。在植物中,程序性减数分裂特异性DNA双链断裂(DSBs)在SPO11复合体的催化下启动重组途径,尽管只有~5%的结果是COs的形成。

近日来自英国的的布里斯托大学的Mark Winfield团队在国际知名期刊Plant Biotechnology Journal杂志上在线发表了名为Identification, characterisation, and rescue of CRISPR/Cas9 generated wheat SPO11-1 mutants的研究论文。为了研究SPO11-1在小麦减数分裂中的作用,该团队使用CRISPR/Cas9对六倍体小麦的所有6个SPO11-1基因拷贝进行编辑。后代的表型显示,所有6个SPO11-1拷贝缺失的植株无法进行染色体联会,缺乏COs,不育。相比之下,携带三种野生型中的任何一种的单拷贝株系在表型上与未编辑的植物在营养生长和育性方面没有区别。此外,我们发现,携带6个经过编辑的带有TaSPO11-1B基因的SPO11-1的小麦突变体的转化,恢复了染色体联会、CO的形成和育性,从而为这种重要农艺作物的修饰重组开辟了一条途径。
在陆地植物中,减数分裂DSB是通过由SPO11-1/SPO11-2异二聚体和MTOPVIB同型二聚体组成的异四聚体形成的。SPO11是一种进化上保守的II型拓扑异构酶。大多数真核生物拥有单一的SPO11拷贝,尽管在一些植物物种中已经发现了多个SPO11的同源基因。例如,拟南芥有三个同源基因,而水稻有五个。在这两种情况下,只有SPO11-1和SPO11-2对减数分裂必不可少。SPO11-1和SPO11-2也在六倍体面包小麦(Triticum aestivum)中被发现,SPO11-2对DSB的形成至关重要。

TaSPO11-1是一个具有15个外显子的复杂基因,该团队专门设计了sgRNA4和sgRNA6对蛋白质活性位点的编码序列(SPO11-1A和SPO11-1D中的Y 129, SPO11-1B中的Y 128)进行分析。

图2:三个亚组同时编辑的SPO11-1植株是不育的
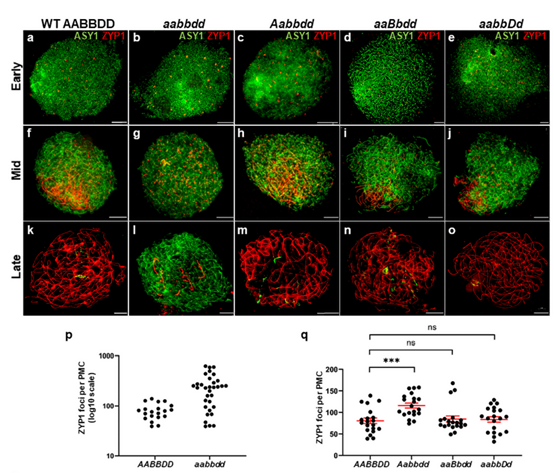
图3:植物细胞学分析
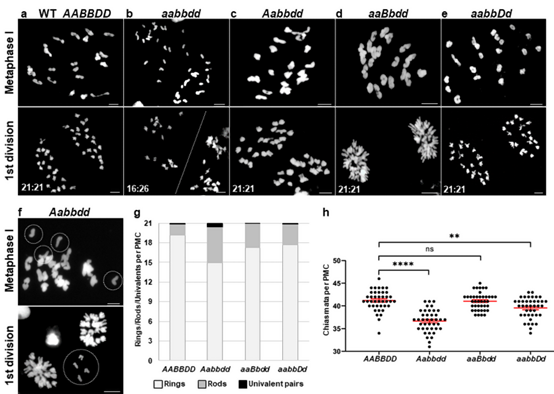
图4:三个亚组同时编辑植株(aabbdd)不能形成交叉


对4株不同程度恢复育性的aabbdd转基因阳性植株进行细胞学分析。
文章来源:植物生物技术Pbj
