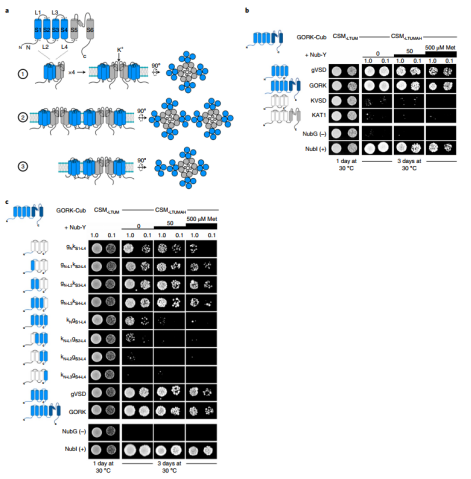
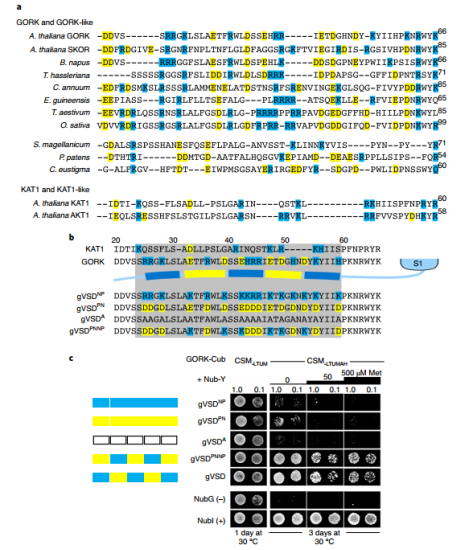
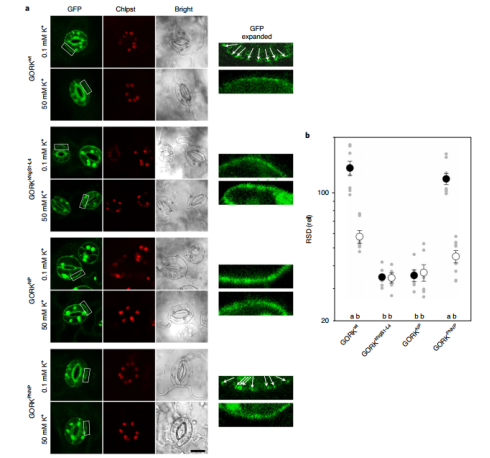
近日,英国格拉斯哥大学植物生理学与生物物理实验室Michael R. Blatt团队在nature plants在线发表了题为“Engineering a K+ channel ‘sensory antenna’ enhances stomatal kinetics, water use efficiency and photosynthesis”的研究论文。该文章确定了带门控的耦合通道聚类机制,并证明了在保卫细胞中原生离子通道工程的潜力,以增强气孔动力学并提高无需碳固定成本的用水效率。气孔在植物叶子的表皮细胞层形成,是CO2从大气扩散到叶肉细胞进行光合作用的主要途径。当打开时,气孔还为从叶片内部空气空间的蒸汽饱和环境到大气的水分损失提供了途径。因此,气孔必须经常平衡光合作用中对CO2的需求与防止叶子干燥的需求。为规避这种碳水平衡带来的一系列不同的挑战,最好通过改变气孔孔径动力学来解决。当减少的H+泵送和Cl−通道的活化相结合以去极化质膜时,气孔关闭,从而促进K+和Cl−从保护细胞中流出。K+流出的主要途径是GORK通道。向外整流通道GORK如拟南芥中密切相关的通道SKOR和向内整流通道 KAT1,属于存在于所有门上的电压门控K+通道的超家族。这些蛋白质形成四聚体的功能通道,每个蛋白质亚基包含六个跨膜α螺旋与胞质N-和C-末端。亚基的前四个α螺旋(S1-S4)形成半自主电压传感器域(VSD),最后两个α螺旋(S5-S6)通过四聚体通道组件中心的膜排列充满水的孔。作者使用基于酵母交配的分裂泛素筛选(mbSUS)来测试截断的通道结构域是否会与全长通道蛋白相互作用,发现了与GORK的相互作用是特定于gVSD结构的。用kVSD的相应结构域在gVSD中进行了顺序结构域替换,以识别gVSD与GORK相互作用的关键区域,结合所必需的主要位点位于GORK通道69个残基的胞质N末端(图1)。(图1 GORK K+通道通过电压传感器域的胞质N端相互作用)进一步,作者将GORK的N端和从芸苔属油菜到小麦和水稻的相关物种通道对齐显示出交替带电区域的模式,利用酵母来测试的残基突变体与全长GORK通道的相互作用。作者得出假设:沿着N端的带电表面的交替使得在寄存器中与GORK的VSD N末端之间的互补带电表面的相互作用成为可能,类似于拉链(图2)。(图2电荷的交替定义了GORK-VSD相互作用)作者利用GORK稳定转化材料和GFP标签观察荧光标记物,通过随后的成像,电生理学和气体交换研究进行。非相互作用的通道突变在拟南芥中稳定表达时抑制GORK聚类(图3)。接下来,作者利用电压钳位下记录来自相同植物的完整保卫细胞的GORK电流,结果证实了每个非相互作用的突变体都减少了外部K+对通道活性的抑制,而GORKPNNP互补挽救了野生型特征。(图3 非相互作用的通道突变在拟南芥中稳定表达时抑制GORK聚类)为了分析GORK突变对气孔动力学和气体交换的影响,作者再次使用用于聚类和通道门控分析的相同植物记录了气体交换特征。使用保护细胞特异性启动子来补充gork空突变体,结果表明N端修饰对GORK的任何影响都应该来自它们对保护细胞K+通量的影响。此外测试了对钾营养的可能影响,结果表明影响GORK门控的突变对K+同源性没有实质性影响,并且与gork空突变体相比,每个互补都足以恢复正常的K+含量。最后作者测试分析了衡量工厂绩效的一种指标WUE和转基因材料的生物重,证实了生物量的增加不是光合作用本身改变的结果。而是补充GORK突变影响其门控,增强气孔动力学以改善WUE并增强碳同化(图4)。(图4 非相互作用的GORK通道突变加速气孔闭合并增强WUE)总之,作者的发现现在证明了改变保护细胞原生离子通道中的门控的功效。通过引入影响GORK通道聚类的突变,改变了K+通量的电压和K+依赖性,增强了保卫细胞膜的转运并加速了气孔运动。这些发现强调了通过“调整”植物原生通道的门控以改善WUE和碳同化而可能获得的好处,从而规避了在保护水的同时确保光合同化生长方面经常相互冲突的需求。文章来源:植物生物技术Pbj