作者首先对热带豆科植物(以脲为转运形式)和温带豆科植物的根瘤转录组数据进行比较。分析发现XMPP、NSH1 和 NSH2只在热带豆科植物的根瘤中上调,GSDA轻微诱导。结合前人的研究,作者创制了XMPP、GSDA、NSH1 和 NSH2 和 XDH 的突变体。探究这些基因是否参与到了热带豆科植株中脲的合成。
在这项研究中,xdh大豆突变体结节积累了大量的黄嘌呤并且不产生脲,同时也检测了IMP的含量与野生型相比突变体中的含量并没有增加说明黄嘌呤主要不是通过 IMP 降解产生的。xmpp 和 xmpp gsda.2 gsda.3 突变体中黄嘌呤积累尿囊素和尿囊酸浓度降低,这表明这些酶参与了脲的生物合成。
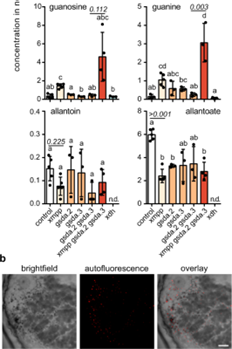
为了评估 NSH1 和 NSH1/NSH2 复合体是否参与在根瘤脲中生物合成,分析了 nsh1 和 nsh2 突变体中根瘤的代谢物含量。由于 NSH2 需要与 NSH1 相互作用才能激活,因此 NSH1的突变体可以被认为是 NSH1 和 NSH2 的功能性敲除。缺乏 NSH1 的根瘤中积累了黄嘌呤,并且尿囊酸含量大大降低,这表明 NSH1 是脲生物合成的关键酶。有趣的是,nsh2根瘤中的代谢物含量与野生型没有区别。虽然这表明 NSH2 不是脲生物合成所必需的,但生化数据显示 NSH1 和 NSH2 相互作用,并且该复合物比单独的 NSH1 具有显着更强的黄嘌呤水解酶活性。
脲的生物合成在受侵染的细胞中开始并最终转移到未侵染的细胞中,但转变点仍不清楚。作者使用启动子-报告系统来研究几种已知参与脲生物合成基因的启动子活性。使用共聚焦荧光显微镜分析在启动子 pXMPP、pGSDA.1、pGSDA.2、pGSDA.3、pNSH1、pNSH2、pXDH、pUOX 或 pALN 控制下表达报告基因的结节的横截面。pXMPP 和三种 pGSDA,在仅在感染细胞中中心区域观察到荧光,这与公共 RNAseq 数据集中来自该基因的相对低丰度的 mRNA 一致。数据表明 XMP 去磷酸化和鸟苷脱氨发生在受感染的结节细胞中。NSH1 和 NSH2 的启动子都仅在与侵染细胞密切相关的未侵染细胞中具有活性。表明,脲的生物合成发生在受感染的细胞中。黄嘌呤在相邻的未感染细胞中被 NSH1/NSH2 复合物水解为核糖和黄嘌呤。作者在与侵染细胞密切相关的未侵染细胞中观察到 UOX 启动子活性。根据这些发现,XDH 在侵染细胞中产生的尿酸盐被转移到侵染区的未侵染细胞中,以通过 UOX 进一步氧化。ALN荧光蛋白定位于内皮层维管束周围的细胞中。
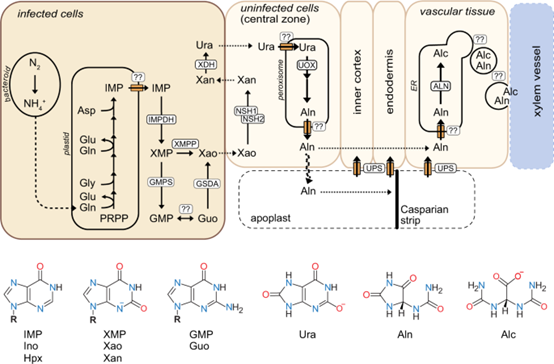
综上所述,作者创制了 XMPP、GSDA、NSH1 和 NSH2 和 XDH 突变体。通过LC-MS 分析了嘌呤核苷酸分解代谢的靶向代谢谱,探究了这些基因在脲生物合成中的作用。此外,探究了这些基因UOX 和 ALN 的细胞表达域。创建了一个根瘤中脲生物合成的模型。
文章来源:植物生物技术Pbj
