再生是生物体生长和生存的一个不可或缺的过程,长期以来一直吸引着生物学家的兴趣。尽管物种间再生的类型和程度差异很大,但植物的全能性使它们成为研究这一显著现象的优越系统。植物再生反应进行分类的一种方法是基于再生模式,即机械损伤诱导的再生和组织培养诱导的再生。机械损伤诱导的再生主要涉及丢失的细胞、组织或器官的替换,而组织培养诱导的再生介导了整个生物体的从头开始重建。其中植物愈伤组织培养是利用植物细胞的全能性,通过器官从头发生的过程产生整个芽/根系统。
芽从头再生是一个由植物生长素和细胞分裂素触发的连续过程,涉及多种植物特异性转录因子(TFs)。只有少数被称为祖细胞的细胞,可以启动从多能性愈伤组织到茎分生组织的旅程,最终产生一个完整的茎系统。在愈伤组织中零星表达的茎干细胞调节因子WUSCHEL (WUS)表明了未来茎分生组织的位置。WUS 和其他促芽因子的这种不均匀表达以及茎分生组织起始的随机性表明愈伤组织具有内在的异质性。一个重要的问题是,特定的愈伤组织细胞是如何被选择为祖细胞的,以及是什么推动它们发展为茎分生组织的?
近日,印度科学教育与研究学院的Kalika Prasad团队在Developmental Cell在线发表了题为“Mechanical conflict caused by a cell-wall-loosening enzyme activates de novo shoot regeneration”的文章,证明了在发育祖细胞中,茎分生组织的再生依赖于极性生长素转运蛋白PINFORMED1 (PIN1)和细胞极性蛋白SOSEKI 2 (SOK2)的定位模式。并进一步表明,祖细胞和相邻细胞之间的机械应力指示细胞极性从头激活茎分生组织形成。
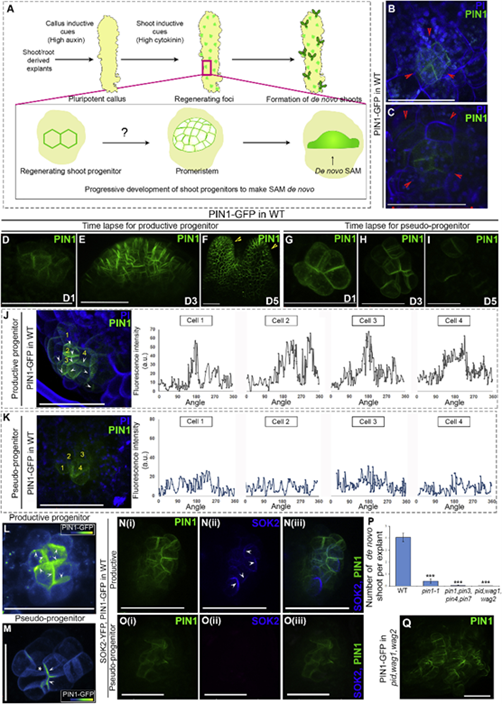
 虽然已知茎分生组织来自愈伤组织的中间层,以响应芽诱导信号,但只有来自该中间层的细胞亚群(称为芽祖细胞)发展成为分生组织。为了研究祖细胞如何发展成分生组织,作者使用PIN1-GFP(已知最早的祖细胞识别标记)标记,并通过共聚焦实时成像技术跟踪它们。虽然能够在一些细胞阶段的细胞膜上检测到 PIN1-GFP,但是观察到并非所有的祖细胞都能发展成分生组织。因此,根据它们的命运将它们分为两种类型:(1)发展成功能分生组织的祖细胞称为“productive progenitors (PPs)”(n, number of PIN1-marked foci = 211/287),(2)中途终止的祖细胞称为“pseudo-progenitors (PSPs)”(n = 76/287)。作者发现,基于 PIN1-GFP 定位模式可以将 PP 与 PSP 区分开来的最早阶段是 6 到 15 细胞阶段。
尽管 PIN1 愈伤组织形成的整个过程中都有表达,但它在祖细胞形成开始期间被限制在特定区域。因此,作者研究了PIN1表达与祖细胞起始阶段的功能相关性。发现PIN1的定位模式与生长素丰度之间存在相关性。总之,上述的研究数据表明,区分 PPs 和 PSPs 的关键特征是细胞极性蛋白的定位模式及其细胞的紧凑排列,而不仅仅是芽祖细胞调控因子的丰度。
图1 Localization pattern of polarity proteins governs the shoot meristem
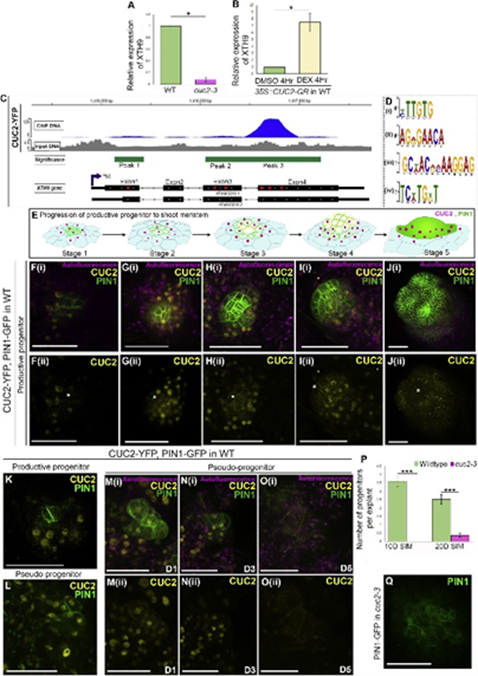
接下来,作者使用比较转录组分析确定了,在再生祖细胞中促进PIN1定位模式的调控因子。观察到非祖细胞中的XTH9(一种细胞壁松弛酶)促进祖细胞生长,而在祖细胞中则相反。并在后续通过在突变体中对XTH9的转录进行分析,发现CUC2(一种促芽因子),能够激活XTH9的表达,从而促进从头分生组织的形成。
图2 CUC2 promotes progenitor progression non-cell autonomously by activating XTH9 in non-progenitor cells
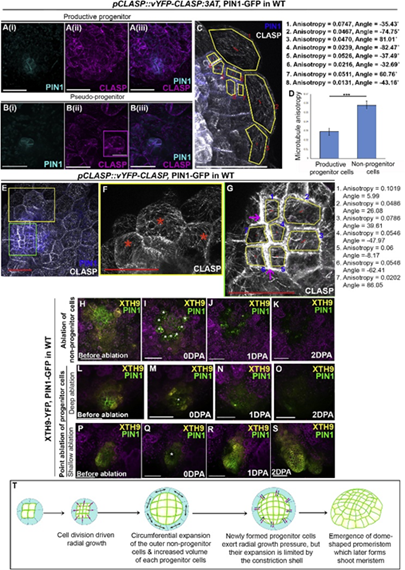
由于XTH9的表达在空间上仅限于非祖细胞的外层,因此作者提出祖细胞的生长速率是否与祖细胞周围的细胞不同。PPs 在 0 h,5 h 30 min,8 h,9 h,和24 h时间点的实时成像显示 PIN1-GFP 标记的祖细胞经历频繁的分裂和细胞扩增,而非祖细胞很少分裂。并在一系列分析之后观察到祖细胞和非祖细胞可能经历了不同的力场。由此研究了物理上干扰这些力是否会影响祖细胞向分生组织的发展。最终结果显示祖细胞周围细胞的遗传或者物理扰动,会损害祖细胞,反之亦然。这些结果说明,在没有组织模式建成的情况下,祖细胞及其周边细胞存在一个反馈回路,作用于芽再生。
图3 De novo shoot meristem formation necessitates mechanical conflict between progenitor and its surrounding cells
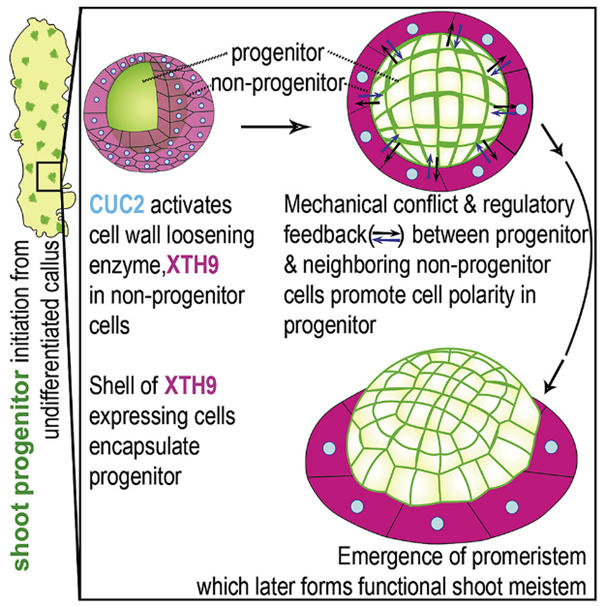
图4 文章总概。极性蛋白的定位模式预测再生祖细胞的命运。
CUC2 仅在非祖细胞的外层中激活细胞壁松弛酶 XTH9。XTH9的空间特异性在祖细胞与其相邻细胞之间产生机械应力。在拟南芥中,祖细胞与其相邻细胞之间的反馈激活了分生组织的形成。
虽然已知茎分生组织来自愈伤组织的中间层,以响应芽诱导信号,但只有来自该中间层的细胞亚群(称为芽祖细胞)发展成为分生组织。为了研究祖细胞如何发展成分生组织,作者使用PIN1-GFP(已知最早的祖细胞识别标记)标记,并通过共聚焦实时成像技术跟踪它们。虽然能够在一些细胞阶段的细胞膜上检测到 PIN1-GFP,但是观察到并非所有的祖细胞都能发展成分生组织。因此,根据它们的命运将它们分为两种类型:(1)发展成功能分生组织的祖细胞称为“productive progenitors (PPs)”(n, number of PIN1-marked foci = 211/287),(2)中途终止的祖细胞称为“pseudo-progenitors (PSPs)”(n = 76/287)。作者发现,基于 PIN1-GFP 定位模式可以将 PP 与 PSP 区分开来的最早阶段是 6 到 15 细胞阶段。
尽管 PIN1 愈伤组织形成的整个过程中都有表达,但它在祖细胞形成开始期间被限制在特定区域。因此,作者研究了PIN1表达与祖细胞起始阶段的功能相关性。发现PIN1的定位模式与生长素丰度之间存在相关性。总之,上述的研究数据表明,区分 PPs 和 PSPs 的关键特征是细胞极性蛋白的定位模式及其细胞的紧凑排列,而不仅仅是芽祖细胞调控因子的丰度。
图1 Localization pattern of polarity proteins governs the shoot meristem
接下来,作者使用比较转录组分析确定了,在再生祖细胞中促进PIN1定位模式的调控因子。观察到非祖细胞中的XTH9(一种细胞壁松弛酶)促进祖细胞生长,而在祖细胞中则相反。并在后续通过在突变体中对XTH9的转录进行分析,发现CUC2(一种促芽因子),能够激活XTH9的表达,从而促进从头分生组织的形成。
图2 CUC2 promotes progenitor progression non-cell autonomously by activating XTH9 in non-progenitor cells
由于XTH9的表达在空间上仅限于非祖细胞的外层,因此作者提出祖细胞的生长速率是否与祖细胞周围的细胞不同。PPs 在 0 h,5 h 30 min,8 h,9 h,和24 h时间点的实时成像显示 PIN1-GFP 标记的祖细胞经历频繁的分裂和细胞扩增,而非祖细胞很少分裂。并在一系列分析之后观察到祖细胞和非祖细胞可能经历了不同的力场。由此研究了物理上干扰这些力是否会影响祖细胞向分生组织的发展。最终结果显示祖细胞周围细胞的遗传或者物理扰动,会损害祖细胞,反之亦然。这些结果说明,在没有组织模式建成的情况下,祖细胞及其周边细胞存在一个反馈回路,作用于芽再生。
图3 De novo shoot meristem formation necessitates mechanical conflict between progenitor and its surrounding cells
图4 文章总概。极性蛋白的定位模式预测再生祖细胞的命运。
CUC2 仅在非祖细胞的外层中激活细胞壁松弛酶 XTH9。XTH9的空间特异性在祖细胞与其相邻细胞之间产生机械应力。在拟南芥中,祖细胞与其相邻细胞之间的反馈激活了分生组织的形成。