在阅读文章之前让我们先来回忆两个知识点:什么是雄性不育?什么是杂种优势?
雄性不育:所谓雄性不育就是在有性繁殖过程中,由于生理上或遗传上的原因造成植物的雌性器官正常,雄性器官不正常,因而不能完成正常授粉的现象。雄性不育是高等植物群体中普遍存在的一种现象。
杂种优势:即杂交种第一代在各方面均比亲本优越的现象,杂交种第一代植株主要表现在具有更快的生长速率和更高的代谢功能,从而导致器官发达、体型增大、产量提高,或者表现在抗病、抗虫、抗逆性、生存力等的提高,因此杂种优势也被广泛用于提高农作物的产量。
目前,杂交育种已经广泛应用于水稻、玉米、高粱、大豆、番茄等作物的生产中,尤其是玉米杂交种率几乎可达100%,这也足以看出杂交种在农业生产中的重要性。然而,遗传分离现象的存在会使得杂交种第一代自交后丧失杂种优势。因此,为了获得稳定的杂交种,核心问题就是要避免自交。在农业生产中采用的解决办法通常是利用雄性不育株构建杂交系。由于雄性不育株的花粉发育不良,只能作为母本接受配套父本的花粉并与之杂交,从而确保产生的子代是具有优势的杂交种。但是稳定雄性不育株的常规筛选不仅耗时而且还很费力,由此产生的高成本也阻碍了作物中杂种优势的应用。近年来,随着基因编辑系统的不断完善,科学家们尝试在不同物种中利用基因编辑技术创制雄性不育系,并相继获得了成功,这大大简化了杂交育种的流程。那么,接下来就和伯小远一起看看在不同物种中利用基因编辑创制雄性不育系的研究进展吧~
水稻
2016年11月22日,庄楚雄团队在Scientific Reports上发表了题为“Development of Commercial Thermo-sensitive Genic Male Sterile Rice Accelerates Hybrid Rice Breeding Using the CRISPR/Cas9-mediated TMS5 Editing System”的研究论文,该团队将CRISPR/Cas9系统应用在雄性不育系的培育中,开发出了新的“无外源基因”TGMS(热敏雄性不育)系,为显著加快不育系的育种周期,促进杂种优势的开发提供了新思路。
作者首先选取了国内最常用的热敏型雄性不育基因TMS5作为靶标基因,为了获得具有高编辑和低脱靶效率的靶序列,作者在TMS5编码区筛选出了10个靶位点,并随机分为5组,将一组中的两个靶序列克隆到同一个载体中,使用CRISPR/Cas9系统进行靶向诱变,结果显示靶向TMS5a和TMS5b位点的双元载体(称为TMS5ab载体)具有最高的靶向编辑效率(图1)和低脱靶效应(表1)。为了获得TGMS系,作者将T0不育植物在低温(~22°C,日平均温度[DAT])下进行处理,从而获得具有恢复生育力的植物,此外,作者还在T1植物中获得了“无外源基因”TGMS系(图2、图3)。
综上所述,作者认为可以利用有效的基因编辑载体TMS5ab,来培育具有潜在应用价值的无外源基因TGMS系。值得关注的是,作者在一年内利用TMS5ab在两个水稻亚种中培育出了11种具有杂交育种应用前景的无外源基因TGMS系。这进一步说明该方法有助于推动杂种优势的开发。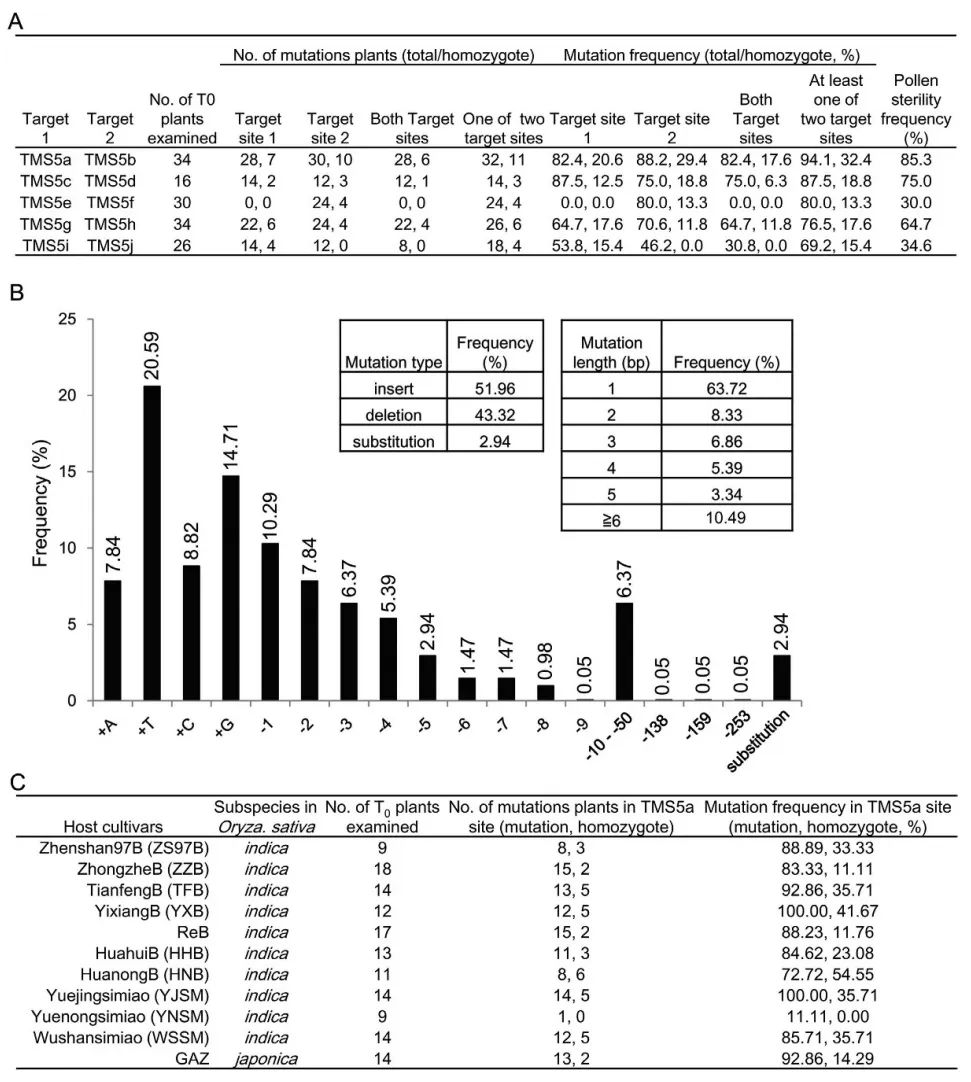 图1 T0植物中的突变类型和频率(Zhou et al., 2016)。(A)T0植物中不同靶点的突变情况和花粉不育频率。(B)CRISPR/Cas9诱导的突变类型和频率。(C)TMS5ab载体在两个不同水稻亚种的11个品种中诱导的突变频率。
图1 T0植物中的突变类型和频率(Zhou et al., 2016)。(A)T0植物中不同靶点的突变情况和花粉不育频率。(B)CRISPR/Cas9诱导的突变类型和频率。(C)TMS5ab载体在两个不同水稻亚种的11个品种中诱导的突变频率。
表1 在TMS5a和TMS5b靶点的假定脱靶位点中检测到突变(Zhou et al., 2016)。
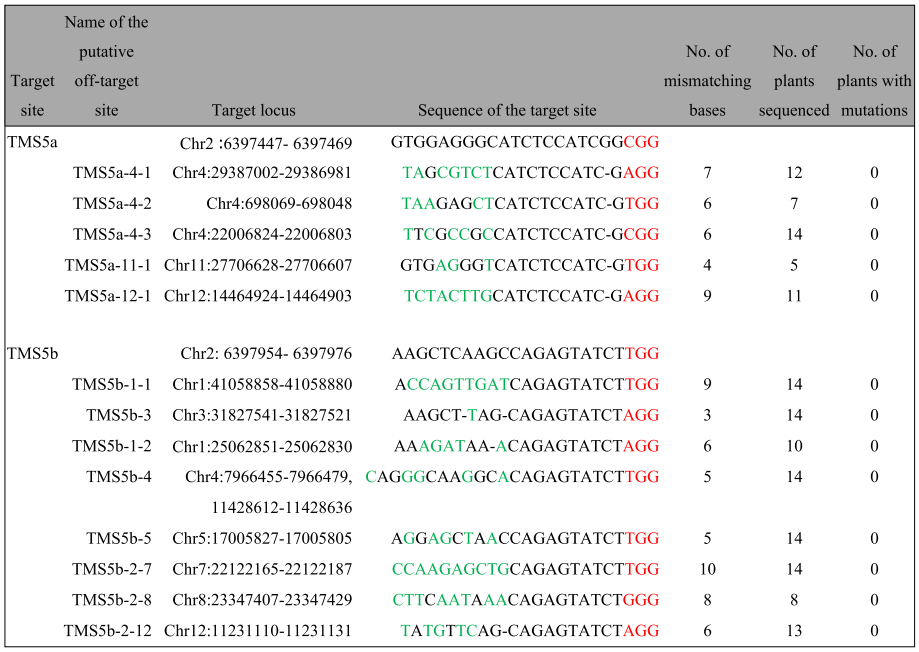
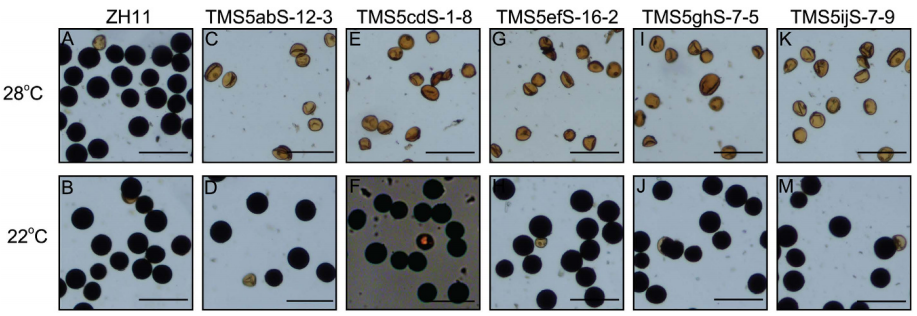
图2 高温和低温下CRISPR/Cas9系统诱导的TGMS系的花粉育性(ZH11背景)(Zhou et al., 2016)。

图3 高温处理的野生型和热敏型雄性不育基因植物的植株形态(Zhou et al., 2016)。
2020年7月8日,谢传晓团队在Molecular Plant上发表了题为“Genome Editing Enables Next-Generation Hybrid Seed Production Technology”的研究论文,在这篇文章中作者利用CRISPR/Cas9的新系统简化了杂交育种的流程,仅需一步就可以创制不育系和保持系,并同时解决了不育基因导入和不育株筛选的问题。
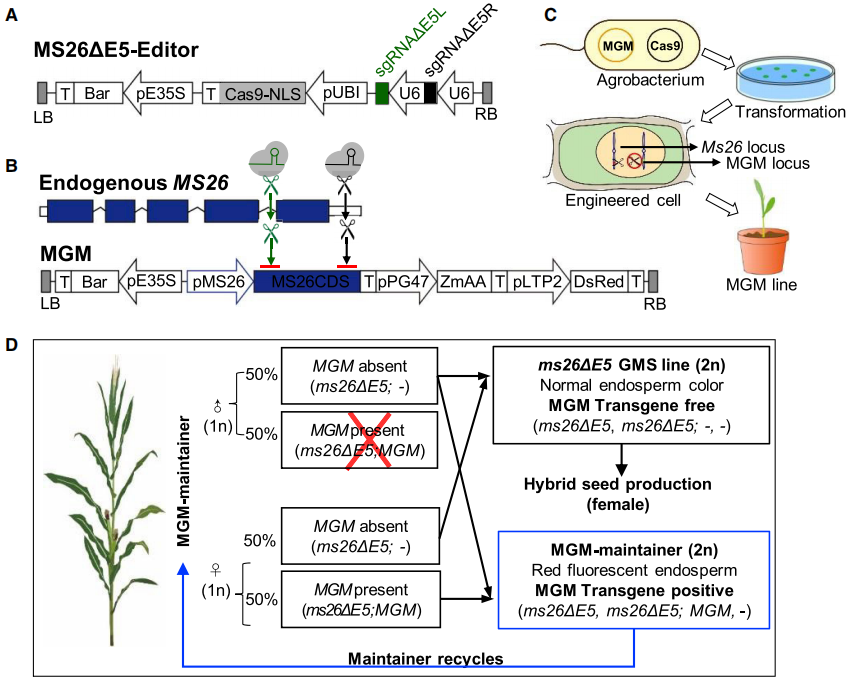 图4 利用基因编辑创制玉米不育系和保持系的载体构建示意图及工作流程(Qi et al., 2020)。(A)MS26ΔE5-Editor载体构建示意图;(B)MGM载体构建示意图;(C)将MS26ΔE5-Editor载体与MGM载体共转植物细胞;(D)在杂交种子生产中生成雄性不育系(GMS)和保持系(MGM)的工作流程,♂:雄性配子体、♀:雌性配子体、1n:单倍体细胞、2n:二倍体细胞。
图4 利用基因编辑创制玉米不育系和保持系的载体构建示意图及工作流程(Qi et al., 2020)。(A)MS26ΔE5-Editor载体构建示意图;(B)MGM载体构建示意图;(C)将MS26ΔE5-Editor载体与MGM载体共转植物细胞;(D)在杂交种子生产中生成雄性不育系(GMS)和保持系(MGM)的工作流程,♂:雄性配子体、♀:雌性配子体、1n:单倍体细胞、2n:二倍体细胞。
作者首先设计了一个MS26ΔE5编辑器(图4A),来对玉米的内源育性基因ZmMS26进行编辑从而创制雄性不育系。在sgRNA的引导下Cas9蛋白通过剪掉ZmMS26的一小段,使该基因失去功能,从而导致玉米雄性不育。在杂交体系中,使用不育系作为母本,就可以与其他父本优势种杂交获得杂交种。
另外,作者还构建了另一个载体MGM用于创制保持系(图4B),该载体包含三个功能模块:一个是用于恢复植株雄性不育的育性的MS26 CDS表达元件、另一个是能导致花粉失去活性的酶、最后一个是会让玉米粒发出红色荧光的标记基因。通过将MGM载体与MS26ΔE5编辑载体共同转化到宿主细胞中后,由于MGM拷贝中的MS26 CDS缺少sgRNA的靶位点,因此并不会被剪切(图4C)。此外,MGM上携带的花粉淀粉酶基因会造成花粉败育,因此只有不携带MGM的花粉可以作为雄配子,雌配子则一半携带MGM,一半不携带MGM(图4D)。当使用保持系自交时,最终会获得一半不育系和一半保持系。而在后期不育系和保持系的筛选中,由于MGM上含有荧光基因,因此保持系的玉米籽粒会发出红色荧光,这极大的简化了后期的筛选流程(图5)。值得注意的是,在这个系统中,不育系植株不携带外源性的MGM基因,因此只要是与非转基因品种杂交产生的后代,就不是转基因作物。
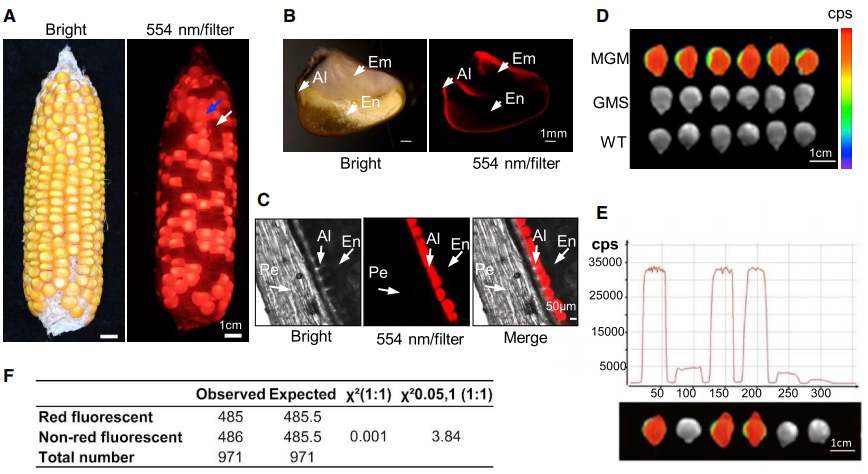
图5 红色荧光标记可作为筛选保持系的标记(Qi et al., 2020)。(A)在明场(左)和554nm波长激发光下用滤光片(右)拍摄的玉米籽粒表型;(B)在体式显微镜(Nikon SMZ1500, Japan)下,携带DsRED的保持系种子纵向剖面的彩色图像;(C)共聚焦显微镜图像显示保持系种子中单层胚乳糊粉中DsRED(红色)的特异性表达;(D-E)使用植物成像系统来区分不同种子中的荧光强度;(F)保持系自交后代分离比的卡方检验结果。
2021年6月2日,冯献忠团队在The Crop Journal上发表了题为“Generation of male-sterile soybean lines with the CRISPR/Cas9 system”的研究论文。该研究表明CRISPR/Cas9技术可在短时间内创制稳定的大豆细胞核雄性不育突变体,加速大豆雄性不育系的育种周期,为大豆杂交育种系统提供新的不育系材料。
绒毡层细胞,即最接近花粉的体细胞层,它可以调节孢子发生和花粉壁发育。ABORTED MICROSPORES(AMS)基因是一种影响拟南芥绒毡层发育的bHLH转录因子。作者利用拟南芥AMS的蛋白序列,鉴定了AMS基因的两个大豆直系同源物Glyma.10G281800和Glyma.20G107500(图6),并分别命名为GmAMS1和GmAMS2,通过CRISPR/Cas9技术对大豆中的这两个AMS同源物进行了靶向编辑来创制稳定的大豆细胞核雄性不育系(图7)。结果显示CRISPR/Cas9技术对GmAMS1的靶向编辑导致大豆产生了雄性不育的表型,与野生型花粉相比,Gmams1-1突变植物的花粉粒呈现出皱缩的形态(图8),而GmAMS2的靶向编辑未能获得雄性不育系。GmAMS1不仅影响花粉壁的形成,还影响小孢子和绒毡层细胞的发育,它可以通过调节细胞程序性死亡(Programmed cell death,PCD)来控制大豆绒毡层的发育和降解(图9)。
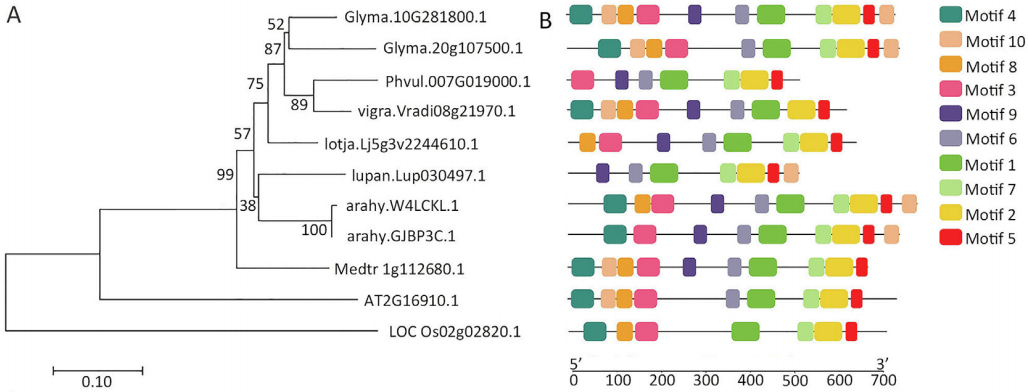 图6 GmAMS的系统发育树及其保守基序分析(Chen et al., 2021)。(A)GmAMS的系统发育树;(B)GmAMS的保守基序及其同源物,由彩色框表示。
图6 GmAMS的系统发育树及其保守基序分析(Chen et al., 2021)。(A)GmAMS的系统发育树;(B)GmAMS的保守基序及其同源物,由彩色框表示。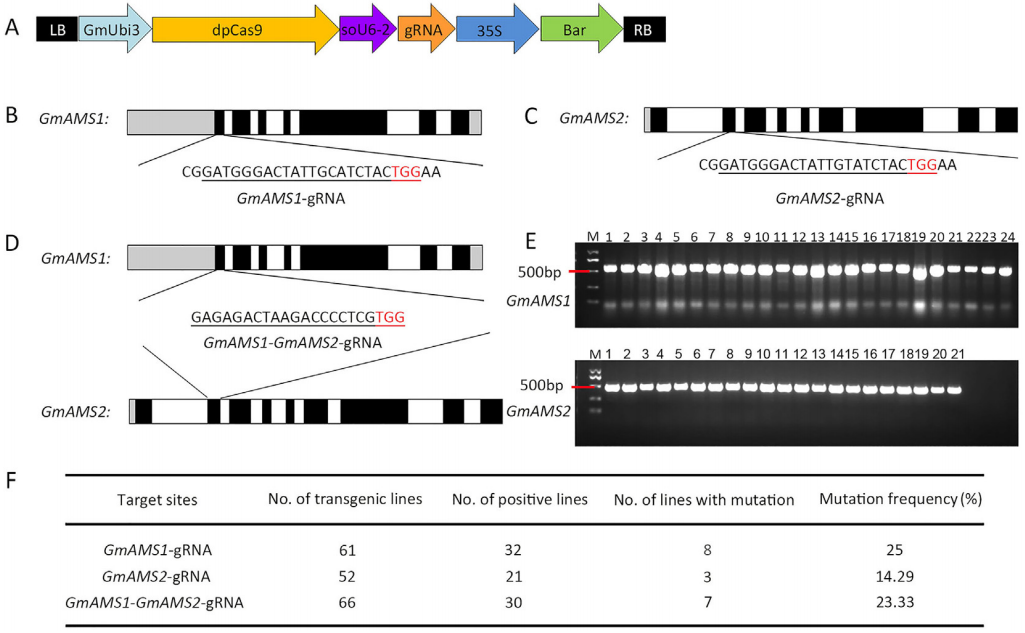 图7 CRISPR/Cas9载体和GmAMS基因中靶位点的结构图(Chen et al., 2021)。
图7 CRISPR/Cas9载体和GmAMS基因中靶位点的结构图(Chen et al., 2021)。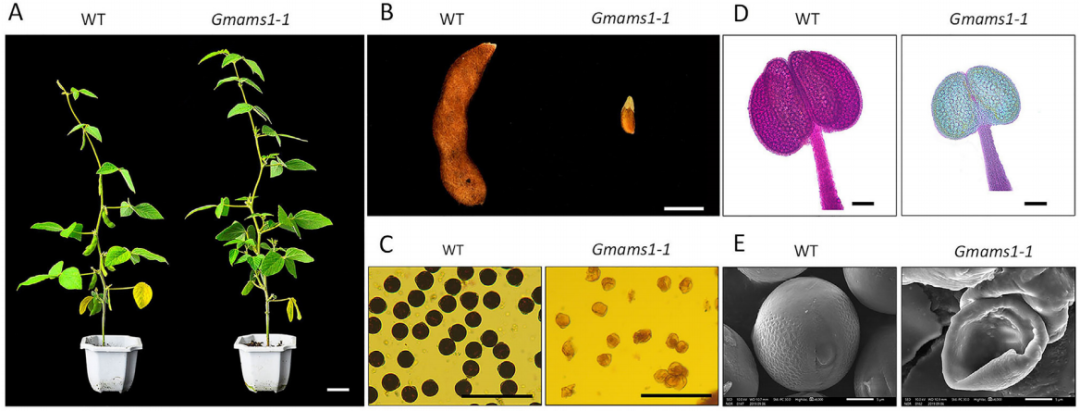 图8 Gmams1-1突变体植物表现出雄性不育(Chen et al., 2021)。(A)WT和Gmams1-1植物的表型对比;(B)WT和Gmams1-1植物成熟时的豆荚对比;(C)WT和Gmams1-1花粉粒的I2-KI染色;(D)WT和Gmams1-1花粉粒的亚历山大染色;(E)WT和Gmams1-1花粉粒的扫描电镜照片。
图8 Gmams1-1突变体植物表现出雄性不育(Chen et al., 2021)。(A)WT和Gmams1-1植物的表型对比;(B)WT和Gmams1-1植物成熟时的豆荚对比;(C)WT和Gmams1-1花粉粒的I2-KI染色;(D)WT和Gmams1-1花粉粒的亚历山大染色;(E)WT和Gmams1-1花粉粒的扫描电镜照片。
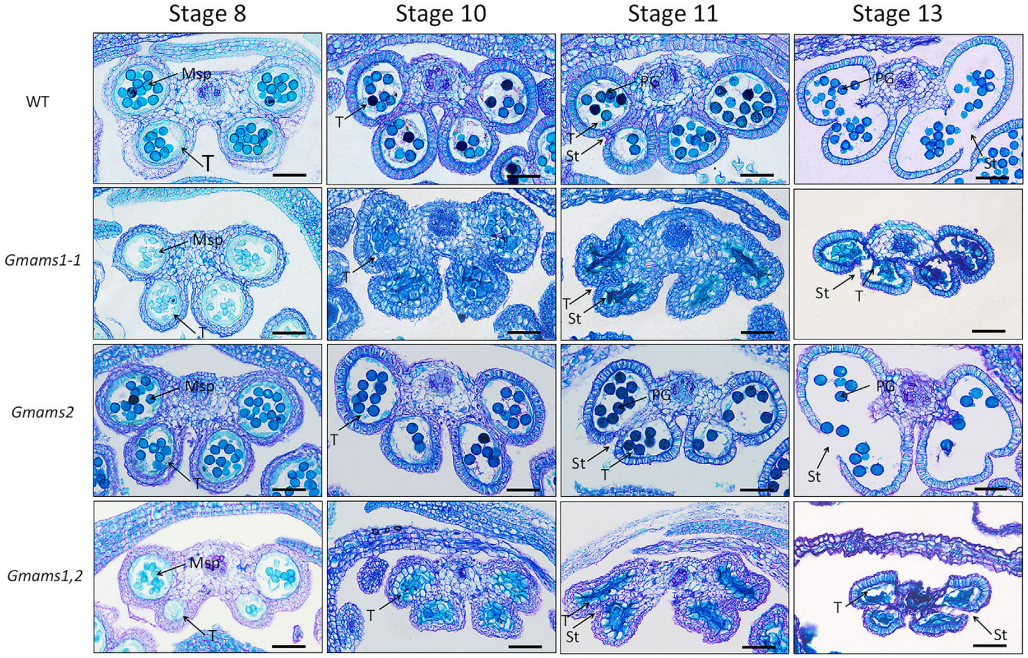 图9 来自WT和Gmams突变体的四个发育阶段的花药裂片的切片,用甲苯胺蓝染色后的结果(Chen et al., 2021)。绒毡层(T)、小孢子母细胞(Msp)、隔膜(St)和花粉粒(PG)由箭头指示。
2020年1月10日,李传友团队和李常保团队合作在The Plant Journal上发表了题为“A biotechnology-based male sterility system for hybrid seed production in tomato”的研究论文。该研究提出了一种利用基因编辑技术在番茄骨干自交系背景中快速创制雄性不育系和保持系,并有效应用于杂交种子生产的策略。
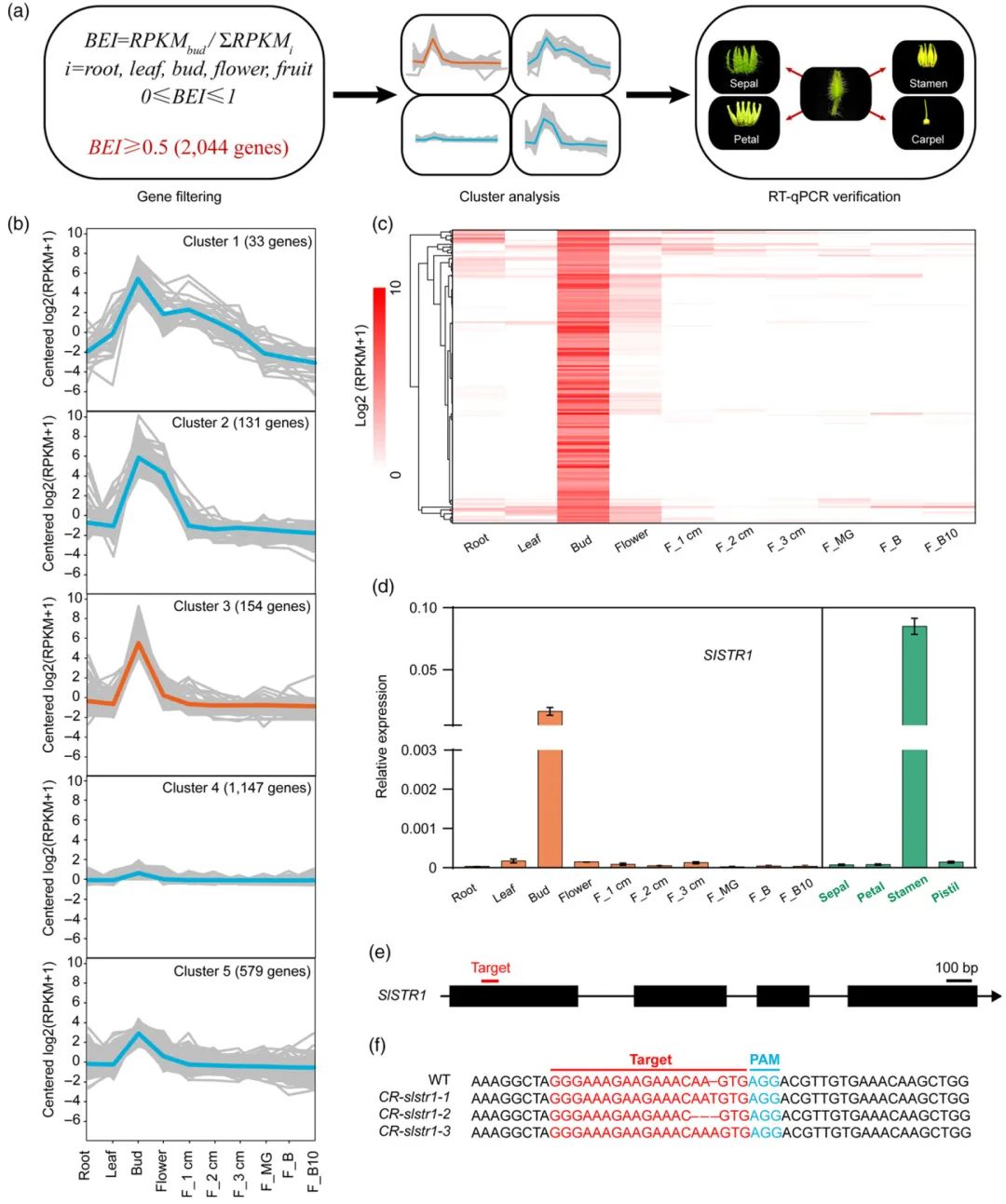
首先为了创制雄性不育系,作者在番茄的基因组中鉴定到了154个在雄蕊中特异性表达的基因,结合数据分析最终选取其中的一个基因SlSTR1作为靶标基因(图10)。在此基础上,作者设计了Cas9/sgRNA载体用于靶向SlSTR1的第一个外显子,通过将Cas9/sgRNA载体转化至番茄骨干自交系TB0993中对SlSTR1进行定向敲除,最终筛选出来了TB0993背景的雄性不育系(图11)。紧接着,作者将正常功能的SlSTR1基因和控制花青素合成的SlANT1基因连锁在一起,共同转回到前期筛选到的雄性不育系中,最终得到了育性恢复的紫色保持系。最后,将雄性不育系作为母本,保持系作为父本进行杂交,其子代会分离出一半非转基因的雄性不育系和一半转基因的保持系,通过幼苗的颜色可以非常方便的区分出雄性不育系(非紫色)来用于杂交育种(图12)。
图10 鉴定在番茄雄蕊中特异性表达的基因(Du et al., 2020)。(a)鉴定在雄蕊中特异性表达基因的方法;(b)划分成五个簇的2044个基因的表达模式;(c)来自簇3的154个基因在不同组织中的表达热图;(d)SlSTR1在不同组织中的表达情况;(e)示意图说明了sgRNA靶向SlSTR1编码序列的位点(红线);(f)从三个CR-slstr1(T0代)株系中鉴定到的代表性突变。
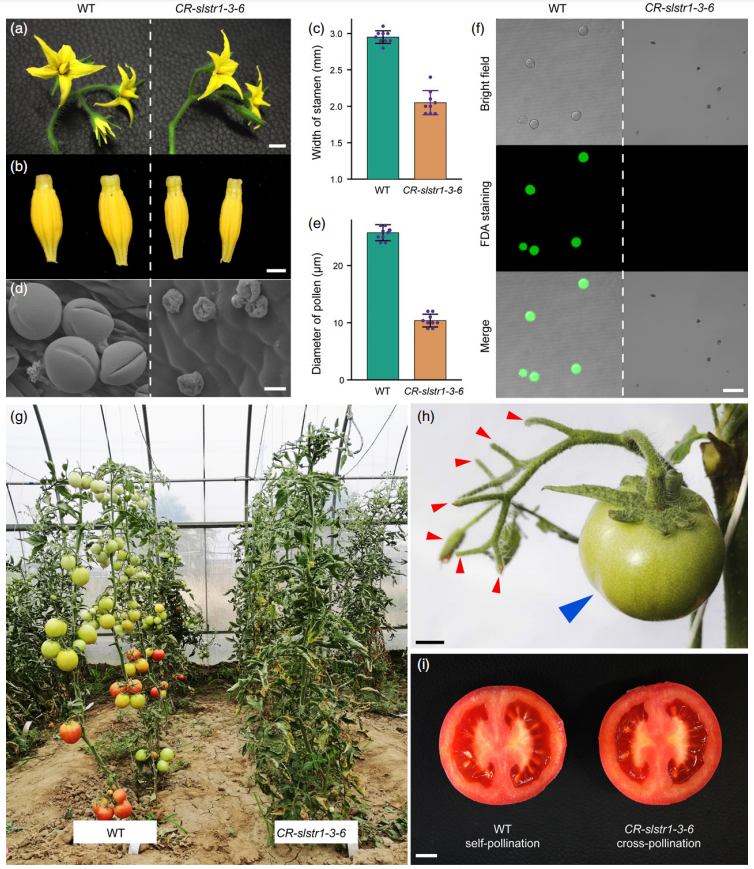
图11 CR-slstr1-3-6植株在花粉发育和雄性生育力方面存在缺陷(Du et al., 2020)。(a-c)野生型(WT)和CR-slstr1-3-6花的形态比较;(d)WT和CR-slstr1-3-6花粉粒的扫描电镜照片;(e)WT和CR-slstr1-3-6花粉粒的直径;(f)WT和CR-slstr1-3-6花粉粒在开裂阶段的活力;(g)田间种植的CR-slstr1-3-6植株的结实缺陷表型照片;(h)CR-slstr1-3-6植物在自花授粉后不结果(红色箭头);(i)CR-slstr1-3-6植物经WT花粉异花授粉后产生带种子的果实。
图9 来自WT和Gmams突变体的四个发育阶段的花药裂片的切片,用甲苯胺蓝染色后的结果(Chen et al., 2021)。绒毡层(T)、小孢子母细胞(Msp)、隔膜(St)和花粉粒(PG)由箭头指示。
2020年1月10日,李传友团队和李常保团队合作在The Plant Journal上发表了题为“A biotechnology-based male sterility system for hybrid seed production in tomato”的研究论文。该研究提出了一种利用基因编辑技术在番茄骨干自交系背景中快速创制雄性不育系和保持系,并有效应用于杂交种子生产的策略。
首先为了创制雄性不育系,作者在番茄的基因组中鉴定到了154个在雄蕊中特异性表达的基因,结合数据分析最终选取其中的一个基因SlSTR1作为靶标基因(图10)。在此基础上,作者设计了Cas9/sgRNA载体用于靶向SlSTR1的第一个外显子,通过将Cas9/sgRNA载体转化至番茄骨干自交系TB0993中对SlSTR1进行定向敲除,最终筛选出来了TB0993背景的雄性不育系(图11)。紧接着,作者将正常功能的SlSTR1基因和控制花青素合成的SlANT1基因连锁在一起,共同转回到前期筛选到的雄性不育系中,最终得到了育性恢复的紫色保持系。最后,将雄性不育系作为母本,保持系作为父本进行杂交,其子代会分离出一半非转基因的雄性不育系和一半转基因的保持系,通过幼苗的颜色可以非常方便的区分出雄性不育系(非紫色)来用于杂交育种(图12)。
图10 鉴定在番茄雄蕊中特异性表达的基因(Du et al., 2020)。(a)鉴定在雄蕊中特异性表达基因的方法;(b)划分成五个簇的2044个基因的表达模式;(c)来自簇3的154个基因在不同组织中的表达热图;(d)SlSTR1在不同组织中的表达情况;(e)示意图说明了sgRNA靶向SlSTR1编码序列的位点(红线);(f)从三个CR-slstr1(T0代)株系中鉴定到的代表性突变。
图11 CR-slstr1-3-6植株在花粉发育和雄性生育力方面存在缺陷(Du et al., 2020)。(a-c)野生型(WT)和CR-slstr1-3-6花的形态比较;(d)WT和CR-slstr1-3-6花粉粒的扫描电镜照片;(e)WT和CR-slstr1-3-6花粉粒的直径;(f)WT和CR-slstr1-3-6花粉粒在开裂阶段的活力;(g)田间种植的CR-slstr1-3-6植株的结实缺陷表型照片;(h)CR-slstr1-3-6植物在自花授粉后不结果(红色箭头);(i)CR-slstr1-3-6植物经WT花粉异花授粉后产生带种子的果实。
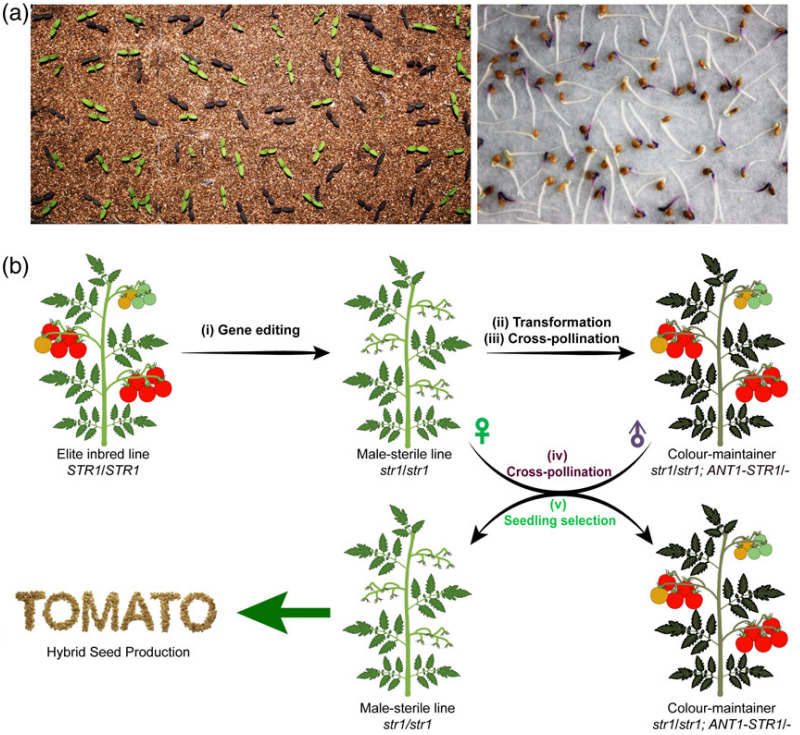 图12 新型番茄雄性不育系统的创制流程(Du et al., 2020)。(a)在幼苗阶段区分雄性不育植物(绿色子叶)和保持系植物(紫色子叶)的代表性照片。(b)基于基因编辑技术的杂交种子生产示意图。
2021年12月28日,王涛、董江丽团队在Plant Biotechnology Journal上发表了题为“Construction of genic male sterility system by CRISPR/Cas9 editing from model legume to alfalfa”的研究论文。该研究建立了利用高效基因编辑工具创制苜蓿隐性核不育系及其保持材料的方法,极大地促进了全球苜蓿产业的杂交优势利用。
作者首先在模式植物蒺藜苜蓿(Medicago truncatula)中通过反向遗传学策略筛选雄性不育靶标基因。前期研究报道称,禾本科中的雄性不育是由编码葡萄糖-甲醇-胆碱(Glucose-Methanol-Choline, GMC)氧化还原酶基因的单基因纯合突变造成的,但是在双子叶植物中尚无相关报道。因此,作者通过对苜蓿、拟南芥、水稻、玉米中的GMC氧化还原酶家族基因进行系统发育分析,发现蒺藜苜蓿Medtr5g011020与已报道的雄性不育基因遗传距离较近,结合该基因在花蕾发育早期雄蕊中特异性高表达,因此推断Medtr5g011020可能参与苜蓿雄性育性调控,并将其命名为MtNP1。接下来,作者利用优化过的苜蓿基因编辑工具靶向野生型中的MtNP1来构建突变材料,筛选到的纯合移码突变体Mtnp1没有观察到任何营养生长缺陷,在生殖阶段表现为只开花未结果的雄性不育表型(图13),因此可以确定MtNP1能作为创制苜蓿雄性不育系的靶标。最后,作者在紫花苜蓿( Medicago sativa L. )栽培种(中国地方品种“保定苜蓿”)中利用优化过的苜蓿基因编辑工具靶向MsNP1来获得突变体材料。结合表型分析发现,全部等位移码突变的Msnp1突变体为雄性不育系材料(图14)。而三等位杂合突变体能够产生成熟花粉粒,可作为保持材料与全等位突变体进行姊妹交,后代能够稳定产生不育系基因型,并筛除外源转基因片段(图15)。
此外,通过重测序发现MsNP1靶点基因组区段在苜蓿全球核心种质中高度保守,因此,只要建立可行的转化体系,该研究设计的雄性不育基因编辑方案适用于不同遗传背景的紫花苜蓿材料。
图12 新型番茄雄性不育系统的创制流程(Du et al., 2020)。(a)在幼苗阶段区分雄性不育植物(绿色子叶)和保持系植物(紫色子叶)的代表性照片。(b)基于基因编辑技术的杂交种子生产示意图。
2021年12月28日,王涛、董江丽团队在Plant Biotechnology Journal上发表了题为“Construction of genic male sterility system by CRISPR/Cas9 editing from model legume to alfalfa”的研究论文。该研究建立了利用高效基因编辑工具创制苜蓿隐性核不育系及其保持材料的方法,极大地促进了全球苜蓿产业的杂交优势利用。
作者首先在模式植物蒺藜苜蓿(Medicago truncatula)中通过反向遗传学策略筛选雄性不育靶标基因。前期研究报道称,禾本科中的雄性不育是由编码葡萄糖-甲醇-胆碱(Glucose-Methanol-Choline, GMC)氧化还原酶基因的单基因纯合突变造成的,但是在双子叶植物中尚无相关报道。因此,作者通过对苜蓿、拟南芥、水稻、玉米中的GMC氧化还原酶家族基因进行系统发育分析,发现蒺藜苜蓿Medtr5g011020与已报道的雄性不育基因遗传距离较近,结合该基因在花蕾发育早期雄蕊中特异性高表达,因此推断Medtr5g011020可能参与苜蓿雄性育性调控,并将其命名为MtNP1。接下来,作者利用优化过的苜蓿基因编辑工具靶向野生型中的MtNP1来构建突变材料,筛选到的纯合移码突变体Mtnp1没有观察到任何营养生长缺陷,在生殖阶段表现为只开花未结果的雄性不育表型(图13),因此可以确定MtNP1能作为创制苜蓿雄性不育系的靶标。最后,作者在紫花苜蓿( Medicago sativa L. )栽培种(中国地方品种“保定苜蓿”)中利用优化过的苜蓿基因编辑工具靶向MsNP1来获得突变体材料。结合表型分析发现,全部等位移码突变的Msnp1突变体为雄性不育系材料(图14)。而三等位杂合突变体能够产生成熟花粉粒,可作为保持材料与全等位突变体进行姊妹交,后代能够稳定产生不育系基因型,并筛除外源转基因片段(图15)。
此外,通过重测序发现MsNP1靶点基因组区段在苜蓿全球核心种质中高度保守,因此,只要建立可行的转化体系,该研究设计的雄性不育基因编辑方案适用于不同遗传背景的紫花苜蓿材料。 图13 蒺藜苜蓿Mtnp1突变体的雄性不育表型(Ye et al., 2021)。(A)对Mtnp1突变体的全株(A1)、花(A2)、雄蕊和雌蕊(A3)、亚历山大染色(A4)、I2-KI染色(A5)进行分析;(B)Mtnp1突变体和野生型之间的人工杂交结果。
图13 蒺藜苜蓿Mtnp1突变体的雄性不育表型(Ye et al., 2021)。(A)对Mtnp1突变体的全株(A1)、花(A2)、雄蕊和雌蕊(A3)、亚历山大染色(A4)、I2-KI染色(A5)进行分析;(B)Mtnp1突变体和野生型之间的人工杂交结果。
图14 紫花苜蓿雄性不育系的表型(Ye et al., 2021)。(A)对Mtnp1突变体的花(A1)、雄蕊和雌蕊(A2)、亚历山大染色(A3)、I2-KI染色(A4)的分析;(B)Mtnp1突变体和野生型之间的人工杂交结果。
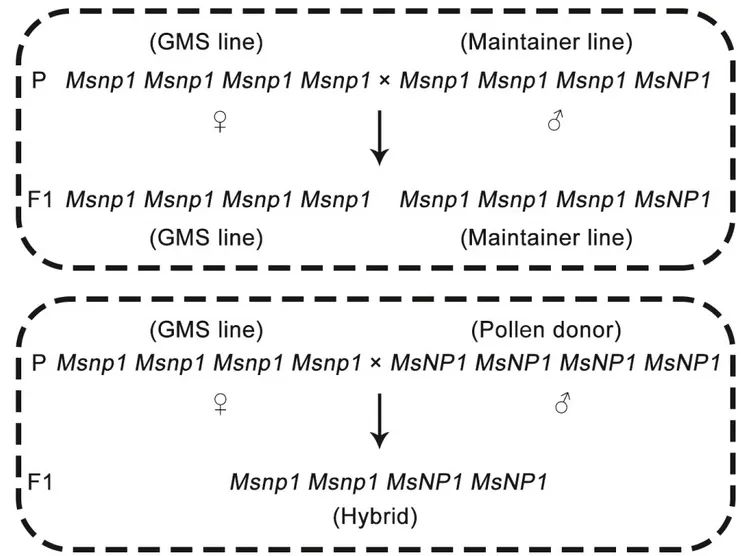
图15 在紫花苜蓿中利用雄性不育系育种的流程图(Ye et al., 2021)。
杂种优势利用是作物遗传改良的重要方式,杂交种创制的核心在于控制授粉,以人工去雄授粉的方式进行杂交制种存在制种成本高、杂交纯度难保证等风险,因此最佳解决方案就是采用雄性不育系(利用基因编辑创制雄性不育系比较适合细胞核隐性雄性不育,对于其它的雄性不育可能不太适合,例如细胞核显性雄性不育、细胞质雄性不育等,原因大家自己去思考哦!)。在本文中小远为大家介绍了在水稻、玉米、大豆、番茄、苜蓿这五种植物中,利用基因编辑技术快速创制雄性不育系的研究进展,这些研究成果为产生雄性不育系提供了一种新颖而方便的策略。当然,除了上述物种外,该方法也在其它物种中也有所应用,但因文章篇幅所限,小远今天就不一一列举了,大家感兴趣的话可以查阅相关资料哦,最后为大家简单总结一下研究思路,希望能帮助大家更好的理解本文!
研究思路: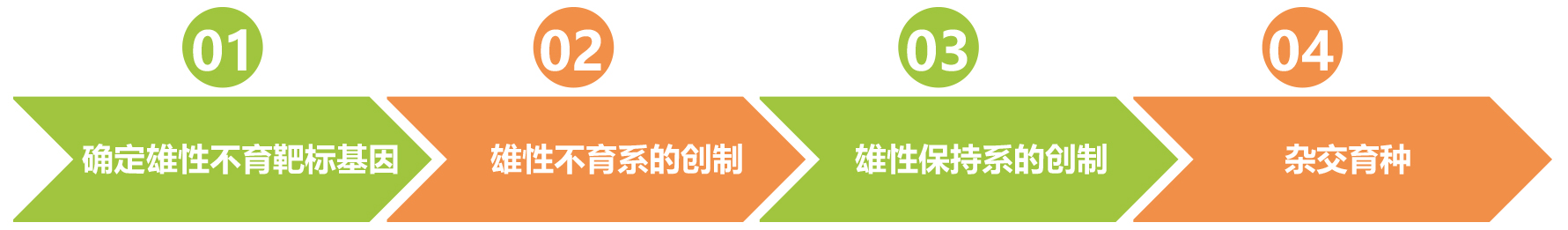 参考文献:
Chen X, Yang SX, Zhang YH, et al. Generation of male-sterile soybean lines with the CRISPR/Cas9 system. The Crop Journal. 2021, 9(6): 1270-1277.
Du M, Zhou K, Liu Y, et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant J. 2020, 102(5): 1090-1100.
Qi X, Zhang C, Zhu J, et al. Genome Editing Enables Next-Generation Hybrid Seed Production Technology. Mol Plant. 2020, 13(9): 1262-1269.
Ye Q, Meng X, Chen H, et al. Construction of genic male sterility system by CRISPR/Cas9 editing from model legume to alfalfa. Plant Biotechnol J. 2021, 20(4): 613-615.
Zhou H, He M, Li J, et al. Development of Commercial Thermo-sensitive Genic Male Sterile Rice Accelerates Hybrid Rice Breeding Using the CRISPR/Cas9-mediated TMS5 Editing System. Sci Rep. 2016, 6: 37395.
参考文献:
Chen X, Yang SX, Zhang YH, et al. Generation of male-sterile soybean lines with the CRISPR/Cas9 system. The Crop Journal. 2021, 9(6): 1270-1277.
Du M, Zhou K, Liu Y, et al. A biotechnology-based male-sterility system for hybrid seed production in tomato. Plant J. 2020, 102(5): 1090-1100.
Qi X, Zhang C, Zhu J, et al. Genome Editing Enables Next-Generation Hybrid Seed Production Technology. Mol Plant. 2020, 13(9): 1262-1269.
Ye Q, Meng X, Chen H, et al. Construction of genic male sterility system by CRISPR/Cas9 editing from model legume to alfalfa. Plant Biotechnol J. 2021, 20(4): 613-615.
Zhou H, He M, Li J, et al. Development of Commercial Thermo-sensitive Genic Male Sterile Rice Accelerates Hybrid Rice Breeding Using the CRISPR/Cas9-mediated TMS5 Editing System. Sci Rep. 2016, 6: 37395.